11 Laks i havet – kunnskapsstatus, forskningsbehov og flaskehalser
Marianne Holm (HI), Lars Petter Hansen (NINA) og Jens Christian Holst (HI)
11.1 Rapportens innhold og avgrensninger
Denne rapporten inneholder en kort oppsummering av dagens kunnskap om postsmolt og laks i havet inkludert ennå upubliserte resultater fra prosjektet «Laks i Norskehavet», en oversikt over årsaker til dødelighet hos laks i det marine miljø, interaksjoner mellom laks og andre fiskearter i havet, en diskusjon om mulighetene for at laks kan bli tatt som bifangst i pelagisk fiske, samt en vurdering av konsekvenser av disse faktorene både for laksen og forvaltningen av laksebestandene. Datagrunnlaget vil bli drøftet, og det vil bli pekt på «flaskehalser» der det er behov for ytterligere innhenting av kunnskap.
Fra merkeforsøk vet vi at de norske laksebestandenes vandringer dekker store deler av det nordlige Atlanterhavet, men denne utredningen begrenser seg til å omhandle kun de områder som anses som primære leveområder for norsk laks, nemlig Nordsjøen, Norskehavet inkludert deler av Grønlandshavet og Barentshavet. Øst- og Vest-Grønland utelates fordi områdene ikke anses som primære leveområder for norsk laks, men der det er det rimelig at dette området trekkes inn i diskusjonen, vil det bli tatt med.
Det er innlysende at med meget korte tidsserier for laks fra det åpne hav, er det på nåværende tidspunkt vanskelig å trekke statistisk helt sikre konklusjoner. Store deler av laksens antatte oppvekstområder er ennå mangelfullt dekket eller ikke undersøkt i det hele tatt. Det foreligger derfor ikke konkrete vitenskapelige beviser for enkelte av de emner som vil bli fremlagt for diskusjon. Mesteparten av materialet er presentert ved møter i Det internasjonale havforskningsråd (ICES) og i Den nordatlantiske laksevernkommisjon (NASCO), men noen, av dataene er ikke endelig bearbeidet. Hypotesene baserer seg på observasjoner gjort i løpet av de første årene med fiskeforsøk i Norskehavet, og vi tar derfor forbehold om at resultatene kan endres når man får lengre tidsserier og en mer fullstendig dekning av området .
11.2 Innledning
Det er generelt lite kunnskap om laksens livshistorie i det marine miljø. Laksesmolten forlater elvene om våren/forsommeren for å komme tilbake etter en–fire år for å gyte. Siden midten av 1970-tallet har det vært en økning i dødeligheten av laks i havet, og dette synes å gjelde generelt i store deler av laksens utbredelsesområde (Anon. 1997). Denne faller sammen med nedgangen i laksefangstene i de samme områdene. Tilbakegangen av laks synes å være størst i Canada. Også i Europa er nedgangen betydelig, særlig for mellom- og storlaks av skotsk opprinnelse, men ikke av samme størrelsesorden som i Canada. I Norge har den generelle nedgangen av laks vært betydelig, men ikke i samme størrelsesorden som i Canada og Skottland (Anon. 1997).
De tidligere undersøkelsene gjort på laks i havet dreide hovedsakelig om voksen fisk ( Rosseland 1971, Hansen et al . 1993, Hansen & Pethon 1985, Anon. 1983), mens undersøkelsene av postsmolt (nylig utvandret smolt de første månedene i havet) – hovedsakelig grunnet metodiske problemer–begrenset seg til utsettings-, vandrings- og predasjonsstudier i fjorder og den nære kyst (Gunnerød et al. 1988; Holm et al. 1982; Hvidsten og Møkkelgjerd 1987; Hvidsten og Lund 1988). Det er også tatt postsmolt som sporadiske bifangster ved Havforskningsinstituttets (HI) undersøkelser av pelagiske fiskearter i åpent hav.
I 1991 ble det fanget postsmolt i partrålforsøk i Trondheimsfjorden (Hvidsten et al. 1992). Det ble også tatt 36 postsmolt som bifangst ved et partråltokt etter sild i overflaten i den nordlige del av Norskehavet. Disse individene var de første postsmolt som er beskrevet fra nordområdet (Holst et al. 1992). HIs Fangstseksjon hadde i begynnelsen på 90-tallet utviklet en pelagisk forsøkstrål, som rigges til å fiske fra overflaten og nedover. Redskapet ble antatt å være velegnet også for fangst av laks, og ble testet i Norskehavet sommeren 1993. Det viste seg å fungere godt til formålet, og det åpnet for en mer systematisk innsamling av postsmolt laks i forbindelse med HIs nystartede Norskehavsprogram (Mare Cognitum). Etter 1994 har lakseundersøkelser vært inkludert i HIs toktprogram, og dette året startet også et aktivt samarbeide med NINA. I begynnelsen på 90-tallet ble det dessuten utført en større nordisk studie med prøvefiske og merking av voksen laks vinterstiden i området rundt Færøyene som antas å være overvintringsområde for europeisk laks (Jacobsen et al. 1996, Anon. 1998). Disse studiene sammen med undersøkelser av vandringer (Jonsson 1993) og vandringsatferd til akustisk merket smolt i Høgsfjorden – Kvitsøy området (Holm et al. 1982 og 1984) og i Trondheimsfjorden (Hvidsten et al. 1995) danner utgangspunktet for den kunnskap man har begynt å etablere om den unge laksens vandringsveier og generelle biologi i den tidlige marine fase. Også data presentert av Arbeidsgruppe for laks (NASWG) ved Det internasjonale havforskningsråd (ICES) og tidsserier fra NINA er brukt som grunnlag.
11.3 Postsmolt i norskehavet og tilgrensende områder
11.3.1 Metoder og materiale
I de fleste tilfeller er det ved HIs sommertokter brukt en s.k. Åkratrål med en åpning på ca. 25 x 25 m som er rigget med ekstra oppdrift i overtelnen, spesielle dører og to store blåser på hver av trålvingene. Trålens overtelne går ved denne riggingen helt i overflaten og redskapet fisker fra 0 til ca. 25 m dyp (Valdemarsen og Misund, 1995). Vanligvis tråles det med en fart på 3–3,5 knop i 30 minutter. Metoden har vist å egne seg godt for fangst av postsmolt, men det antas at eldre fisk i stor grad unnslipper pga. for lav tauehastighet. Vanligvis tråles det med 30–60 nautiske mils mellomrom i transekter med en avstand på 60 -100 nm. De aktuelle toktene har hovedsakelig startet i sør og gått nordover. Hovedartene som undersøkes av HI på disse toktene, er de viktige kommersielle bestandene sild, kolmule og makrell, men det taes også prøver flere andre arter. I forbindelse med trålstasjonene blir det tatt salt- og temperaturprofiler og det blir samlet inn plankton med ulike redskaper samt gjort meteorologiske observasjoner. Det har av økonomiske årsaker ikke vært mulig for HI å gjennomføre havgående tokt rettet spesielt mot lakseundersøkelser, og dataene som presenteres er samlet inn ved at eget personell er satt ombord i fartøyene for spesielt å registrere og opparbeide fangster av laks. Dette innebærer at stasjonsnettet ikke er optimalt med hensyn til dekning av lakseforekomstene.
11.3.2 Kunnskapsstatus – postsmolt
Smoltutvandringen fra elvene i Norge viser en stor variasjon og foregår i et tidsrom fra slutten av april til begynnelsen av august avhengig av temperaturen i elven og i sjøen utenfor. Fisken har da tilbragt fra 1–5 år i ferskvann, avhengig av temperatur og fødeforhold i elven. Den yngste smolten produseres fra elvene i sør, mens bre-elver og nordlige elver produserer eldre smolt. Observasjoner av akustisk merket smolt på utvandring, sonarobservasjoner ved havbeiteutsettinger og partrålforsøk ved kysten (Holm et al . 1982, 1991 og 1997, Hvidsten et al . 1992 og 1995) viser især at smolten under gitte hydrografiske forhold vandrer ut i små stimer i overflaten, og kan forflytte seg meget raskt ved å utnytte utadgående overflatestrømmer.
11.3.3 Fordeling, antatte spredningsruter i havet, påvirkning fra havstrømmer
I 1991–1998 er det i tidsrommet mai–august blitt gjort over 500 trålhal i overflaten i Norskehavet. Fangstmønsteret viser at postsmoltens vandringer vest av Shetland og i Norskehavet må være sterkt knyttet til overflatestrømmene. Postsmolten følger først kyststrømmene, siden de sterke strømmene langs kontinentalskråningene og til slutt strømmene ute i det åpne Norskehavet. Også disse strømmene er knyttet til strukturer på bunnen, som kanten av Vøringplatået. Vel ute i Norskehavet ser det ut til at postsmolten spres utover i beiteområdet.
En slik vandringsrute kan eksemplifiseres med det vi nå vet om den irske postsmolten. Det er blitt fanget mye postsmolt i området langs Wyville-Thompson ryggen mellom Shetland og Færøyene både på norske og skotske tokt. På grunnlag av et større antall gjenfangster av «snute»-merker (kodete og magnetiserte mikromerker injisert i laksesnuten) i området er det vist at en vesentlig del av denne fisken er irsk. Langs kontinetalsokkelen vest av Shetland går en sterk nordøstlig strøm som utenfor Møre møter den norske kyststrømmen. Strømmene deler seg mot nord i flere komponenter, noe som også synes å reflekteres i fangstmønsteret av postmolt som går ut i en «vifteform» lenger nord. På grunnlag av gode kunnskaper om strømsystemene, analyser av elvealdersfordeling i fangstene av postsmolt og merkegjenfangster kan vi i dag med stor sannsynlighet si at den irske, skotske og engelske postsmolten holder en rute nordover i Norskhavet som ligner mye på den som vi antar at også norsk postsmolt fra elver sør for Stadt holder. Postsmolt fra norske elver nord for Stadt vil muligens følge kyststrømmen nordover langs norskekysten og komme ut i havet lengre nord og øst enn den sørnorske, irske, engelske og skotske postsmolten. På denne måten fordeles postsmolten til ulike områder, og forskjellige faktorer vil påvirke de ulike bestandenes liv i havet.
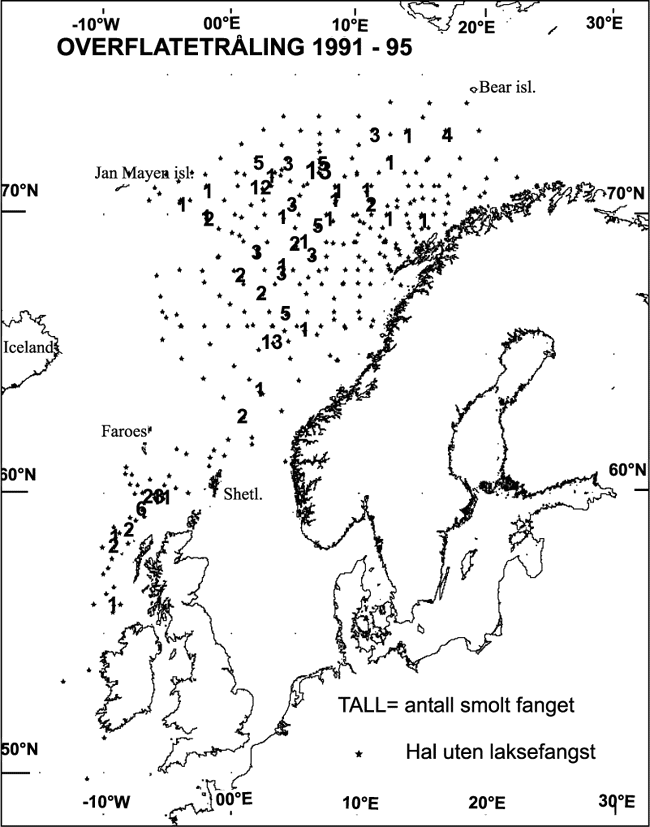
Figur 11.1 Trålhal med og uten laksefangster i 1991–95: Tokt med F/F J. Hjort og G.O. Sars i juni– august.
Kilde: HI.
For postsmolt fra Finmark har vi så langt ingen konkrete kunnskaper, men i forhold til de generelle kunnskaper vi har ervervet om postsmoltens vandringer i havet så langt, kan det settes opp en hypotese: Basert på de strømforhold og temperaturer som opptrer i Barentshavet, forventer vi vandring mot klokken ut av Barentshavet, dvs. først en nordlig til nordøstlig vandring og siden en vestlig vandring sørover til vinteren. Barentshavet er for kaldt for laks om vinteren, og laksen fra dette området overvintrer med stor sannsynlighet i Norskehavet, noe som er bekreftet ved merkeforsøkene ved Færøyene (Anon. 1998).
Vi har registrert at postsmolten er blitt fanget i områder med høy salinitet som karakteriserer det Atlantiske vannet i Norskehavet, i motsetning til kystvannet som har lavere saltholdighet. Lignende er også observert i en skotsk undersøkelse (Shelton et al. 1996 og 1997). Vi tolker dette dithen at postsmolten har sine primære beiteområder i det atlantiske vannet. Fra den geografiske fordelingen av fiskeriene i havet på 60- og 70-tallet vet vi at ved høyere alder trenger laksen inn i kaldere og mindre salt vann med større innblanding av arktisk vann, og beiter da nærmere opptil eller i polarfronten.
11.3.4 Alder, størrelse og opphav
Gjennomsnittsstørrelser hos postsmolt fanget på ulike tokt i 1995-97 viser at smoltstørrelsen øker nordover (Figur 11.2). Dette kan ha sammenheng med at fisken har vært lenger i sjøen og har hatt en lengre beiteperiode når den blir fanget i nord, men det kan også bero på at de nordlige områdene er blitt undersøkt på et senere tidspunkt på sommeren. Størrelsesavhengig vandring, dvs. at de største fiskene vandrer fortest og lengst i forhold til utgangspunktet, er beskrevet som en generell egenskap hos pelagisk fisk. Det er rimelig at dette også gjelder for laks. Basert på alder ved utvandring til sjøen (avlest fra otolitter og skjell) må en betydelig del av den laks som er fanget i det nordlige Norskehavet, være fra de sørlige delene av laksens utbredelsesområde (fra sørvest Norge og sørover), ettersom innslaget av én- og toårssmolt er dominerende (Figur 11.3). I disse områdene går fisken ut allerede i april. I midt- og nordnorske elver forekommer ingen énårssmolt og nesten ingen toårssmolt. Få treårssmolt og nesten ingen fireåringer er blitt registrert i fangstene (unntatt i 1991). Det kan derfor se ut som om vi ikke har vært tilstrekkelig langt nord eller nordøst for å treffe oppholdsstedene til nordnorsk og russisk fisk under sommertoktene.
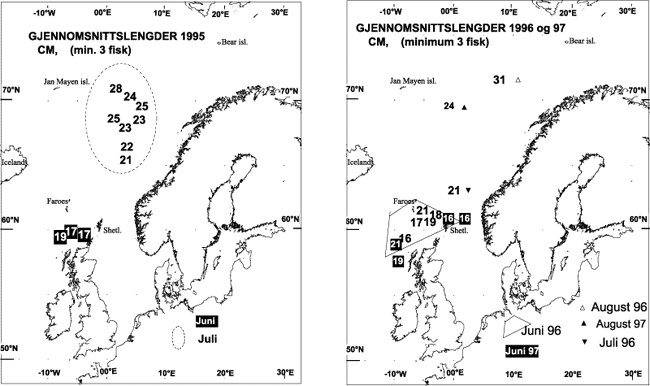
Figur 11.2 Gjennomsnittslengder (cm) for postsmolt i juni og juli 1995 (venstre) og juni, juli og august 1996–97 (høyre). Gjennomsnitt beregnet fra stasjoner med minimum tre fisk i fangsten.
Kilde: HI.
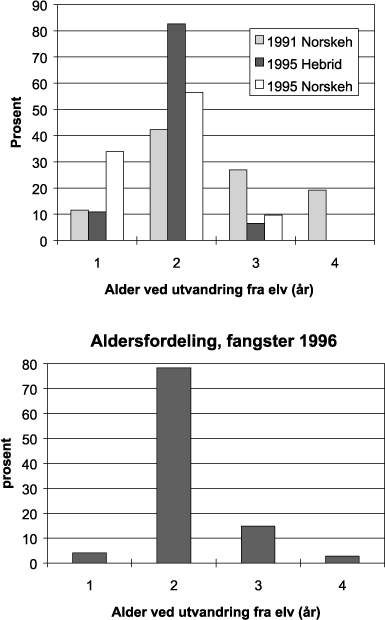
Figur 11.3 Alder ved utvandring fra elv hos trålfanget postsmolt og laks i Norskehavet med tilgrensende områder i 1991–95 (øvre figur) og i 1996 (nedre figur). Avlest fra otolitter og skjell.
Kilde: HI.
11.3.5 Innslag av oppdrettet fisk
Ved aldersavlesningen har vi registrert skjell av postsmolt med vekstsoner i ferskvannsfasen som tyder på at fisken har vært i kultur. Disse fiskene kan imidlertid både være rømt fra oppdrett, og laks som er satt ut for kultivering av vassdrag. Laks som har vært i oppdrett, og som har rømt på et senere stadium, kan derimot lett gjenkjennes også ved hjelp av ytre karakterer. I fangstene av én- og tosjøvinterfisk har vi registrert ca. 15 % at totalt 25 eldre laks fanget i årene 1991–97 var rømt oppdrettsfisk. Flere av disse ble fanget langt nord og vest i havet, og en av disse nær 73°N, 7°E sammen med villfisk. Ved Færøyene er ca. 25 % av bestanden oppdrettslaks. Dette viser at mange av de rømlinger fra oppdrett som overlever, vandrer ut av fjordene og finnes i de samme havområdene som villfisken.
11.3.6 Merkeregistreringer
Det er inntil 1998 registrert fire mikromerkete postsmolt fra de norske trålfangstene. Én var sluppet i april/mai i elven Test i Sør-England (95), den andre var av irsk opphav (96), mens de to siste merkene (97) var merket og satt ut i henholdsvis den irske Ballywynich-elven og en elv i Wales (navn ikke oppgitt). De to første fiskene ble tatt i slutten av juli nord for den 70. breddegrad, vel 2 000 km nord for utsettingsstedet, mens de to siste ble tatt ved Wyville-Thompson ryggen i Nordsjøen. I Norge foretas det for tiden ikke mikromerking av laks, kun Carlin-merking. Det er ikke registrert Carlinmerker i åpent hav fra HIs tråltokter.
11.3.7 Vurdering av årsaken til observert nedgang i fangstene til havs
Det er sannsynlig at bakevjer i strømgyrer kan danne områder der planktonorganismene konsentreres, og at fisken vil prøve å holde seg i områder med gunstige temperaturer og god mattilgang. Det er også sannsynlig at disse områdene kan variere i omfang og lokalisering fra år til år avhengig av klimatiske forhold. Rapporter fra mange elver om gode fangster av ensjøvinter laks i 1998 støtter antagelsen at dette kan være én årsak til nedgangen i fangstene i 1997. Toktene har hovedsakelig gått fra sør mot nord dvs. i smoltens antatte vandringsretning, og det kan muligens ha medført at vi ved de rutinemessige, pelagiske toktene i årene etter 1995 har ligget «på etterskudd» i henhold til hovedkonsentrasjonene av smolt på nordlig vandring
11.3.8 Pelagiske fiskerier i laksens utbredelsesområde
Det drives fiske etter makrell med flytetrål i internasjonalt farvann i Norskehavet. Fisket foregår til en stor del i områder som grenser mot norsk økonomisk sone og som har vist seg å ha et betydelig innslag av postsmolt laks i perioden juni- august. Videre fiskes det med not og trål etter sild i Norskehavet og etter sild og makrell i Nordsjøen, både innenfor de enkelte tilgrensende lands fiskerisoner og i internasjonalt farvann.. I Barentshavet fiskes det i perioder etter lodde med pelagiske redskaper.
Silde- og loddebestandene viser store svingninger i bestandsutvikling og fangst, mens makrellen har vært mer stabil. Mens sildefisket forventes å reduseres noe i kommende år vil makrellfisket trolig holde seg på nåværende nivå eller øke noe, og loddefisket forventes å starte opp igjen i 1999 eller i 2000.
11.3.9 Vurdering av pelagiske redskaper og bifangst som dødelighetsfaktor for postsmolt
Redskaper som overflatetrål og snurpenot som fisker aktivt i de aller øverste vannlagene, kan ta postsmolt som bifangst. Det ser ut til at laksen, også i åpent hav, hovedsakelig oppholder seg nær overflaten, sannsynligvis i små stimer. Vi har indisier på slik atferd fra våre tokt, der man ofte får flere fisk av gangen, og fordi man først begynte å få laks i fangstene når man begynte å tråle i de øverste 25 m. Store fangster av postsmolt ved skotske trålforsøk i peker også på at disse redskapene kan være effektive hvis de brukes i områder der laksen konsentrerer seg under vandring og beiting. I 1997 fikk vi 142 smolt i et eneste havtimes hal nordøst for Shetland.
Vi har per dags dato for lite data både om ungfiskens oppholdssteder og redskapenes effektivitet til å kunne forta en faglig forsvarlig vurdering av hvilken betydning de pelagiske fiskeriene har for dødelighet hos postsmolt og eldre laks. Likevel ser vi at enkelte pelagiske fiskerier skjer midt i viktige vandringsruter for postsmolten, med det potensiale dette har for økt dødelighet i postsmoltbestandene. Det vil være viktig å øke kunnskapene om disse forhold, men det har vist seg å være vanskelig å få til systematiske undersøkelser av dette.
11.4 Forholdet mellom dødelighet hos laks og miljøforhold i havet
Det er store variasjoner i innsiget av laks i forskjellige år, og det er både langsiktige og kortsiktige svingninger. Dødeligheten av laksen i havet er relativt stor, og det er stor enighet om at de fleste laksene dør tidlig i sjøfasen, på postsmoltstadiet i løpet av de første månedene i havet. Tidligere har man hatt svært dårlige forutsetninger for å forutsi en smoltårsklasses skjebne, og laksen har derfor blitt forvaltet basert på historiske data, spesielt har historisk fangststatistikk blitt brukt som indikator på innsigets størrelse. Resultatet av dette har vært at nedgangsperioder for laksen har blitt dokumentert først mange år etter at de startet. For eksempel var norske laksebestander sterke på midten av 1970-tallet, men tilbakegangen ble ikke fastslått før i begynnelsen av 1980-tallet. Dette resulterte høyst sannsynlig i overbeskatning av en rekke laksebestander i flere år før tiltak for å redusere fangsttrykket ble satt i verk.
11.4.1 Generelt om marin dødelighet
Det er alminnelig antatt at den viktigste naturlige dødelighetsfaktoren i havet er predasjon. (Wheeler & Gardner, 1974). Selv om det er beskrevet en rekke sykdommer og parasitter på laksefisk, er lite dokumentert om hvilken rolle disse spiller for den naturlige dødelighet av laks i saltvann (Håstein & Lindstad 1991). Imidlertid har det blitt observert at lakselus kan påvirke både atferd og dødelighet hos sjøørret, og i Irland er det som et resultat av slik infeksjon observert en betydelig dødelighet av ørret (Whelan 1993). Også i Norge er det observert ørret med store infeksjoner (Anon. 1993), og nyere undersøkelser (Finstad et al. 1994) tyder på at lakselusa har hatt negativ effekt på ørretbestanden i Nordsandvassdraget i Vesterålen. Under et samarbeidstokt mellom Skottland og Norge sommeren 1997 i Nordsjøen og vest av Shetland ble det fanget noen postsmolt med mer enn 40 chalimuslarver av lakselus. Hvis disse henger på fisken til de har oppnådd preadultstadiet, er dette høyt over de infeksjonsrater som fører til dødelighet i forsøk (Grimnes & Jakobsen 1996).
Det kan tenkes at overlevelse av smolt kan variere med tilgjengelighet av mat, spesielt i den første tiden etter utvandring fra ferskvann. For eksempel har Farmer et al . (1978) vist at rett etter at smolten har kommet seg ut i saltvann, øker energiforbruket. Hvis det da er lite mat tilgjengelig, er det på sikt rimelig å anta økt dødelighet. Dessuten, hvis det er lite mat, vil laksen vokse dårligere og følgelig være mer utsatt for predasjon, fordi predasjonen er størrelsesselektiv.
Tidspunktet som smolten vandrer ut i sjøen på har meget stor betydning for overlevelsen. Dette er vist gjennom en rekke utsettingsforsøk av smolt i Inndalselva i Sverige som renner ut i Østersjøen (Larsson 1977). I Imsa er det optimale tidspunkt i mai. I forsøk hvor vi overførte smolt som var foret opp på Ims til sjøvann, merket den og satte ut grupper med jevne mellomrom, fant vi at gruppen som overlevde best var den som ble satt ut på den normale tiden for smoltutvandring. Fisk som ble satt ut senere på sommeren, overlevde meget dårlig, til tross for at denne var større (Hansen & Jonsson 1989). Disse forsøkene støtter hypotesen om at det er et smalt smoltutvandringsvindu om våren/forsommeren som gradvis lukkes utover sommeren.
Det har vist seg at høy vannføring når smolten vandrer ut, kan øke overlevelsen. Dette er observert både i Gaula og i Surna (Hvidsten & Hansen 1988) og i vassdrag i Maine i USA (Hosmer et al. 1979). Årsakene til den økte overlevelsen kan være flere; for eksempel at ferskvannslaget i estuariet og fjorden er såpass tykt at det for eksempel er vanskelig for torsk å trenge gjennom for å ta smolten som befinner seg i det øverste vannlag. Dessuten kan økt turbiditet som skyldes flom medføre at predatorer ser smolten dårligere. En annen mulighet er at høy vannføring og derved høy vannhastighet, kan bringe smolten lengre ut i fjorden på kort tid og derved redusere predasjonen.
Miljøet i havet har stor betydning for en smoltårsklasses skjebne. Temperaturen i sjøen ved utvandring har blitt vurdert til å være svært viktig for overlevelsen av postsmolt (se Ritter 1989). Friedland et al. (1993) har publisert et arbeide som tyder på at arealet av postsmolt habitatet definert etter laksens optimaltemperatur har stor betydning for overlevelsen. Dessuten er miljøforholdene i havet også med på å bestemme vekst og kjønnsmodning hos laksen (Scarnecchia 1984).
Mennesket bidrar også til en betydelig dødelighet av laks i sjøen. Den viktigste faktoren er høsting. Laksen har i Norge i svært mange år vært meget hardt beskattet, i 1980 årene er det estimert at over 90 % av innsiget til enkelte bestander ble fisket opp (Hansen 1988). Etter at betydelige reguleringer på laksefisket ble innført i 1989, har fiskepresset minket, men det er tegn på at dette i de siste par årene igjen har øket noe.
Andre dødelighetsfaktorer kan være lokal forurensing av forskjellig slag, som sammen med annen menneskelig aktivitet som f.eks. fiske, samt naturlige forhold kan gi økologiske forandringer som påvirker laksebestandene. Ofte er det kompliserte sammenhenger mellom en rekke faktorer, og kunnskapen om hvordan dette fungerer er meget liten. I den senere tid har det blitt påvist at en del av dødeligheten i sjøen kan også skyldes forhold laksen blir utsatt for i ferskvann på smoltstadiet. Staurnes et al. (1996) viste at laksesmolt eksponert til surt vann i en kort periode hadde dårligere sjøvannstoleranse enn normalt, og dette ble også bekreftet ved analyse av utsettingsforsøk med laksesmolt i en sur og en kalket elv.
11.4.2 Korrelasjoner mellom miljøvariabler og overlevelse
For å undersøke om det finnes muligheter for generell forutsigbarhet av de forskjellige smoltårsklassers skjebne, er det viktig å undersøke graden av samvariasjon i marin overlevelse mellom forskjellige laksepopulasjoner, med andre ord: har den marine dødelighet for forskjellige laksepopulasjoner de samme årsaker? En første enkel og grov måte kan være å se på om fangstene av laks i forskjellige områder er korrelert. I ICES arbeidsgruppe for laks har man undersøkt om det er sammenheng mellom rapporterte fangster i perioden 1960 til 1993 fra noen av de viktigste lakseproduserende land i Atlanteren, og det er en forbausende stor grad av sammenheng. Alle korrelasjonskoeffisienter er signifikante bortsett fra mellom Norge og England/Wales. Vi vil ikke her gå inn på en detaljert analyse av alle usikkerhetene ved bruk av laksestatistikkene, men til tross for alle feil og mangler ved disse, antyder denne analysen at det er betydelige felles faktorer som bidrar til å regulere overlevelsen av laks i havet.
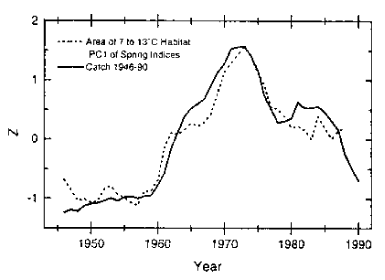
Figur 11.4 Areal av 7–13° om våren sammenholdt med fangster av laks 1960–1990 (uttrykt i «Prinsipal komponent poeng Z)
Kilde: HI.
Friedland et al. (1993) har publisert et arbeide som tyder på at arealet av postsmolt habitatet definert etter laksens antatte optimaltemperatur har stor betydning for overlevelsen. De indikerte at nedgangen i laksebestandene de senere år skyldes at havet hadde blitt kaldere.(Fig.11.4). En foreløpig analyse av overlevelse av smolt fra Figgjoelva viste en høy grad av samvariasjon med Friedlands et al. (1993) postsmolt areal (Hansen og Friedland 1995). Det er interessant at langtidsserien fra Figgjo viser økende overlevelse fram til 1973 og senere en nedadgående trend, noe som er i samsvar med trender i postsmolt arealet.
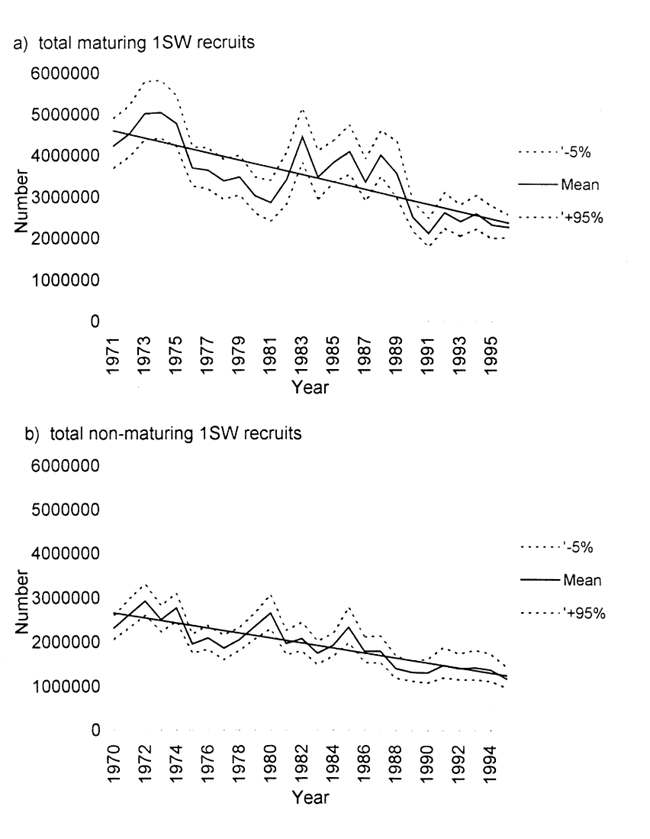
Figur 11.5 Maksimum og minimum estimater av «prefishery abundance» av a) énsjøvinter og b) flersjøvinterlaks i Øst-Atlanteren.
Kilde: HI.
En tredje metode som nå også utprøves på data for Øst-Atlanteren er å bygge opp laksebestanden til mengde fisk før den beskattes (prefishery abundance), etter mønster fra en lignende aktivitet i Vest-Atlanteren (Anon. 1997). Imidlertid er bestandssituasjon mye enklere i Vest-Atlanteren enn i Øst-Atlanteren, men vi skal her kort skissere problemstillingen. Man justerer fangststatistikken av laks i de respektive land for fangst, så bruker man de data man har på totale beskatningsrater i hjemlige farvann, korrigerer for naturlig dødelighet og på den måten får man bygd opp bestanden. Det som er viktig er at man bruker antall laks av forskjellig sjøalder, og ikke vekt. Det er tydelige nedadgående trender både for 1 sjøvinter laks (1 SW) og flersjøvinter laks (MSW) i Øst-Atlanteren (Figur 11.5) Hovedproblemet med denne metoden er alle usikkerhetene med fangststatistikk og at estimering av beskatningsrater kun er basert på få datakilder. Denne metoden er under videreutvikling i ICES, og senere vil dataserien bli sammenlignet med miljøvariabler som for eksempel temperatur.
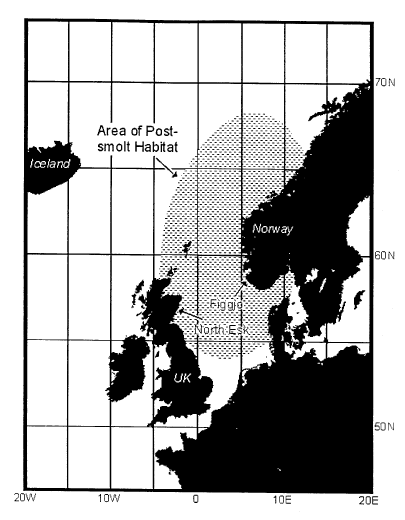
Figur 11.6 Beliggenhet av elvene North Esk og Figgjo Det er antatt at smolt fra de to elvene oppholder seg i det skraverte området de første 1–2 månedene i havet.
Kilde: HI.
En fjerde måte å tilnærme seg denne problemstillingen er å undersøke om overlevelse av laks fra forskjellige indekselver og med forskjellig sjøalder er korrelert. Dessverre har vi ikke så mange indekselver med gode langtidsdata, men gjennom et samarbeid med Skottland og USA har vi sammenlignet de to eneste langtidsserier i Øst-Atlanteren av merket villsmolt siden 1960, tallet, nemlig Figgjoelva på Jæren og River North Esk i Skottland (Friedland et al. 1998). Det interessante ved å sammenligne disse er at de respektive vassdrag renner ut på hver sin side av Nordsjøen, og laksen fra disse blir derfor sannsynligvis eksponert til det samme marine miljø, i hvert fall i den viktige postsmolt perioden (Figur 11.6).
I begge disse elvene har det blitt merket vill smolt siden 1965. Korrelasjonsmatrisen (Anon. 1997) for gjenfangstprosentene av laks som har vært i havet henholdsvis en og to år fra begge bestander, og viser signifikant korrelasjon for alle parametrene. Det er derfor en betydelig grad av samvariasjon i gjenfangstrate mellom North Esk og Figgjo, men der er også vel verdt å merke seg at det er signifikant sammenheng mellom én- og tosjøvinter laks i hvert av vassdragene. Her åpnes derfor muligheten for å kunne forutsi innsiget av tosjøvinter laks på bakgrunn av innsiget av énsjøvinter laks.
Samvariasjonen mellom marin overlevelse og estimert areal med forskjellige overflatetemperaturer i postsmoltområdet som laksen blir eksponert til er blitt undersøkt, og en rekke signifikante sammenhenger er blitt avdekket. Det generelle bildet var svært entydig: Hvis det er kaldt i sjøen i mai (da smolten fra disse to elvene vandrer ut i sjøen) er overlevelsen dårlig. Dette kom sterkest til uttrykk for laks fra Figgjo hvor overlevelsen av 1SW laks var signifikant negative korrelert med arealet av vann med overflatetemperatur på 5–7o C (r = – 0.70; P<0.001). Hvis det er varmt i sjøen i mai, er overlevelsen høy, og dette kom sterkest til uttrykk for laks fra North Esk. For 1SW laks ble det funnet en signifikant positiv korrelasjon mellom overlevelse og arealet av vann med overflatetemperatur på 8–10o C (r = 0.64, P<0.001
11.5 Interaksjoner mellom laks, predatorer, andre fiskebestander og byttedyrbestander
For Norskehavet finnes det gode kunnskaper om de viktigste fiskeartenes bestandsutvikling og økologi. Den store bestanden av norsk vårgytende sild som har beitet i området fra rundt 1988 har medført en stor internasjonal forskninginnsats som har pågått i området på hele nittitallet. Det forgikk også store forskningsprosjekter i området på 50- og 60- tallet da sildebestanden siste gang benyttet Norskehavet til beiteområde. Også i Nordsjøen og Barentshavet legges det ned en stor forskninginnsats fra mange land, og det finnes gode kunnskaper om de viktige fiskeartene og deres utvikling i den aktuelle perioden.
Vi vil i dette avsnittet prøve å vurdere i hvilken grad den negative utvikling som er observert i mange laksebestander i perioden etter 1970 kan skyldes økologiske interaksjoner som predasjon og/eller næringskonkurranse mellom laks og andre arter, eller om klimatiske- eller fiskeridrevne kollapser i vitale byttedyrbestander kan ha påvirket utviklingen. Disse forhold er i liten grad studert direkte for laks, men gitt de kunnskaper som eksisterer om arter og bestander som er aktuelle å dra inn i analysen, kan problemstillingene tilnærmes.
11.5.1 Predatorer
Predatorbestandene som beskatter den norske laksen i havet kan deles i tre hovedgrupper: Fugl, fisk og sjøpattedyr. Vi skal i det etterfølgende foreta en systematisk vurdering av tilgjengelig kunnskap om de tre gruppenes betydning som predatorer for norske laksebestander.
11.5.1.1 Fugl
I alminnelighet vil predasjon fra fugl være sterkest i kystnære områder og avta raskt med avstanden fra land bl.a. fordi mange av de aktuelle predatorartene har en begrenset aksjonsradius. Postsmolten vil videre være særlig sårbar for predatorangrep i løpet av de ca. 48 timene før den er fullt saltvannstilpasset (Handeland et al . 1996). Predasjon fra fugl som beiter langt til havs vil avta i løpet av laksens første år i sjøen fordi laksens raske vekst gjør den for stor som byttedyr for fugl i løpet av denne perioden
Sterk predasjon fra fugl som måker, terner, skarv og fiskeender forekommer nær elveutløpene. Det finnes få studier fra havet, men havsule er trukket frem som en predator på postsmolt i åpne havområder. Det er rimelig å anta at de store konsentrasjonene av postsmolt som opptrer i elveutløpene i utvandringsperioden virker spesielt tiltrekkende på ulike predatorbestander, og at det her skjer en kraftigere predasjon over en kort periode enn i noe annet område i sjøen.
Fugl synes lokalt å kunne gjøre sterke innhugg under smoltutvandringen, men vi vurderer det likevel som usannsynlig at fugl skal spille en vesentlig rolle i den generelle nedgang som er observert i mange laksebestander de senere år. Et viktig argument for dette er at overlevelse også for stor laks er gått kraftig ned etter 1970. Det er utelukket at beiting fra fugl kan forklare nedgang i overlevelse av flersjøvinter laks.
11.5.1.2 Fisk
Den aktuelle perioden for predasjon fra fisk på laks er stort sett knyttet til laksens første leveår år i havet. Med få unntak vil laksen etter dette stort sett være for stor som byttedyr for de fleste aktuelle fiskepredatorene.
Innenfor kategorien fiskepredasjon må også kannibalisme inkluderes. Kannibalisme hos fisk er vanlig både hos typiske planktonetere, som norsk vårgytende sild, og hos fiskespisende fisk, eksempelvis norsk arktisk torsk. I begge de nevnte bestander er kannibalisme beskrevet som en viktig årklasseregulerende faktor.
Ved kysten vurderes følgende arter som potensielle for sterk beiting på postsmolt: torsk, sei, lyr, laks og makrellstørje. Med unntak av lyr må de samme artene regnes som potensielle laksepredatorer i havet.
Det finnes i tillegg en rekke andre arter som også er beskrevet å spise laks, f.eks. ørret og enkelte haiarter. Disse artene omtales ikke spesielt, men dekkes inn i den generelle konklusjonen om fisk som predator.
11.5.1.2.1 Torsk
Lokalekysttorskebestander finnes langs hele norskekysten og i de fleste fjorder. Torsk er beskrevet som predator på laks i nærheten av elveutløp og i fjorder, hvor den er observert å kunne spise store mengder utvandrende laksesmolt (Hvidsten & Møkkelgjerd 1987; Hvidsten & Lund 1988). Det synes som om de lokale torskebestandene har vært i tilbakegang, blant annet er det i Nord-Norge pekt på at de lokale gytebestandene er forsvunnet fra enkelte fjorder.
De viktigste havbestandene av torsk i norske havområder finnes i Nordsjøen og i Barentshavet. Bestanden av torsk i Nordsjøen har vist en negativ utvikling fra tidlig på åttitallet (Anon. 1997), mens Barentshavsbestanden har svingt opp og ned i perioder fra 1950 (Anon. 1997). I Norskehavet finnes torsk i randområdene nær land (Norskekysten, Islands nordkyst og rundt Færøyene) og på sokkelen i Barentshavet mellom Norskekysten og Svalbard. I det åpne hav med dybder på over 600–700 meter finnes torsk kun unntaksvis. Norsk arktisk torsk oppsøker Norskekysten i forbindelse med gyting, men dette faller ikke sammen med smoltutvandringen i tid.
I Havforskningsinstituttets magedatabase på norsk arktisk torsk fra Barentshavet som inneholder i størrelsesorden 80 000 mager er ikke laks observert som byttedyr for torsk. Postsmolten antas å tilbringe det meste av tiden nær overflaten, og de to artene vil derfor ha relativt liten grad av romlig overlapp i åpent hav, noe som magedataene bekrefter. Postsmolt i disse områdene må derfor utgjøre en meget marginal komponent i torskens diett.
Basert på observasjonene ovenfor vurderes det som meget usannsynlig at de oceane torskebestandene kan være av direkte betydning for den negative utviklingen i de norske laksebestandene, og det kan heller ikke påvises en generell bestandsoppgang som skulle tilsi at lokale torskestammer skulle kunne ha avgjørende negativ innvirkning på bestandsutviklingen av norsk laks.
11.5.1.2.2 Sei og lyr
Sei er på linje med torsk beskrevet som en effektiv predator på smolt i elveutløp og fjorder. Seien er med stor sannsynlighet også en effektiv predator på postsmolt i fjordsystemer og i havet, uten at dette er dokumentert. Seibestanden var omtrent dobbelt så store tidlig på syttitallet som i dag (Anon. 1997), og det er observert at seimengdene langs kysten har avtatt i samme periode.
Lyr er i begrenset grad beskrevet som predator på postsmolt, men må tas i betraktning i denne forbindelse. Den beiter pelagisk i fjordene og kystnært og er en typisk fiskespiser. Ved havbeiteforsøk på vestsiden av Sotra i Hordaland i 1991-93 var lyr den desidert viktigste predatoren på smolten (Skilbrei et al. 1998). Bestanden av lyr er liten sammenlignet med sei og torsk, og det drives ikke systematiske bestandsstudier på arten. Det er derfor vanskelig å uttale seg om trender i bestandsutvikling i den aktuelle perioden. Artens utbredelse er begrenset nordover og arten er ikke vanlig nord for Lofoten.
Seibestandens negative utvikling siden syttitallet gjør det lite sannsynlig at sei kan ha hatt avgjørende betydning for de norske laksebestandene etter 1970. Lyrbestandens lave nivå, og artens begrensete utbredelse sannsynliggjør ikke at arten skal ha stor betydning for utviklingen i de norske laksebestandene. Derimot kan lyr antas å være en viktig predator på et lokalt nivå ved kysten og nær elver i Sør Norge.
11.5.1.2.3 Andre fiskepredatorer i åpent hav
Makrellstørje ble observert å spise stor laks langs kysten under størjefiskeriene på 1950- og 1960-tallet (Johannes Hamre, Havforskningsinstituttet, pers. medd ). Størjebestanden var stor på femtitallet men avtok utover sekstitallet. Nedgangen fortsatte på sytti- og åttitallet, og i dag må størja karakteriseres som meget sjelden i norske farvann.
Laks er observert å predatere på postsmolt i fjordsystemene og det er rapportert beiting på postsmolt av laks i havet ved Island (Sturlaugsson pers. medd ). Kannibalisme vurderes å kunne være av betydning for dannelse av årsklassestyrke hos laks, spesielt i år som følger etter sterke årsklasser.
Bortfallet av størjebestanden ansees som positivt for laksebestandene i den aktuelle perioden og har ført til lavere beitepress på lakseinnsiget fra denne arten. Bestanden av voksen laks har generelt vært nedadgående i den aktuelle perioden, og det synes ikke rimelig at kannibalisme kan forklare den negative utviklingen i laksebestandene.
11.5.1.3 Sjøpattedyr og vurdering av predasjon
Norskehavet er rikt på ulike arter sjøpattedyr, og flere av disse er aktuelle som predator på laks. Det er god dokumentasjon på at forskjellige selarter spiser laks, spesielt nær kysten (Pierce et al. 1991, Shearer 1992). I Skottland har det også blitt observert at hvalarten tumler fanger laks (Hislop & Shelton 1993). Når disse og annen vitenskapelig dokumentasjon kombineres med førstehånds enkeltobservasjoner, et bilde som tyder på at de fleste artene av sjøpattedyr i Norskehavet er aktuelle predatorer på laks, kanskje bortsett fra helt spesialiserte planktonetere som blåhval. Det finnes stadig bedre bestandsestimater for ulike sjøpatterdyrsbestander, noe som henger sammen med en stor innsats på dette feltet de siste 10 år. Flere av bestandene synes å være i fremgang etter at fangsten er blitt kraftig redusert eller har stoppet helt. Nyere undersøkelser indikerer bl.a. at bestandsnivåene av grønlandssel i Barentshavet er mer enn dobbelt så stor som tidligere beregnet, og bestanden er estimert til rundt 2.1 millioner dyr i 1998 (Anon. 1999).
Selarter som grønlandsel og klappmyss og hvalarter som vågehval, finnhval og spekkhogger har en utbredelse som totalt sett overlapper med beiteområdene til laksens alle aldersgrupper i havet. Hvis det er slik at sjøpattedyrbestandene har øket etter at fangsten på de fleste av artene har opphørt, kan nedgangen i laksebestandene ha sammenheng med økt beitepress fra sjøpattedyr i laksens beiteområder.
11.6 Konkurranse mellom laks og andre fiskebestander
Under den første delen av sjøoppholdet kan postsmolten sammenlignes med en ordinær planktoneter som sild eller makrell med hensyn til økologisk rolle. Etter hvert som postsmolten vokser endres rollen mot topp-predatorens.
I Norskehavet, som regnes som det viktigste beiteområdet for norsk postsmolt, finnes tre store pelagiske fiskebestander: sild, makrell og kolmule. Silda og makrellen er typisk pelagiske arter som for en stor del beiter i de øvre vannmasser og på denne måten overlapper med postsmoltens vertikale utbredelse. Kolmulen derimot er en mesopelagisk art som beiter på dyp mellom 300–600 meter. Dette betyr at kolmulen og postsmolten i liten grad overlapper vertikalt, men de overlapper delvis i næringsvalg fordi næringsdyrene har stor vertikal utbredelse.
11.6.1 Betydning av byttedyrkonkurranse for laksebestandene
Det nærmeste en kan komme å vurdere om det er konkurranse er å se på en periode da en av de potensielt konkurrerende artene var ute av systemet, og sammenligne denne med at bestanden tok seg opp igjen. En slik situasjon oppstod da bestanden av norsk vårgytende sild kollapset på slutten sekstitallet. Silda er den største pelagiske bestanden i Norskehavet og er derfor et god egnet eksempel fordi dens bortfall og gjenvekst, må ha hatt dramatiske konsekvenser for økologien i hele det nordøstlige Atlanterhav. Silda var borte fra Norskehavet fra 1970 til 1987/1988 og kan ikke ha representert noen konkurrent til de andre pelagiske artene i denne perioden. Ser vi på utviklingen til laksebestandene i samme periode, er det påfallende at man opplevde gode lakseår da silda var borte. I dag er sildebestanden stor, og laksebestandene små. En skal være forsiktig med å trekke den konklusjon at sildas bortgang åpnet for fremvekst av laksen. Sild er velegnet føde både for liten og stor laks, og det er mulig at et viktig byttedyr for laksen forsvant med silda. Det er likevel interessant å trekke paralleller til andre systemer der fremvekst av en art kan åpne for andre. Den generelle betraktning kan gis at dersom silda er en konkurrent til laksen, vil dette kunne slå sterkest ut på laksens vekst i kalde perioder med lav produktivitet og lite mat. I perioder med god produktivitet vil konkurransen ha mindre betydning. For utviklingen i laksebestandene vil dette bety at en fortsatt kan få store årsklasser i de gunstigste årene, mens små årsklasser i kalde år vil reduseres relativt mer. Dersom reell næringskonkuranse, mellom laks og sild i uproduktive perioder må det forventes lavere produktivitet i laksebestandene i fremtiden fordi sildebestanden kan forventes å ligge på et nivå mellom 4 og 10 millioner tonn både på kort og lang sikt.
11.6.2 Laks i havet og byttedyrbestandene
Et stort flertall av de byttedyrstudier som er foretatt på laks i havet viser at laksen er en utpreget opportunist som utnytter de byttedyrarter som til enhver tid er tilgjengelig. I magen på samme laks kan man finne en blanding av fisk, gonatus (blekksprut) og krepsdyr. Likevel er det vist at når fisk av passelig størrelse er tilstede fortrekker laksen disse fremfor andre byttedyr (Hansen & Pethon 1985; Holst et al . 1996).
At laksen er opportunist, betyr at den vil være mindre påvirket av kollapser i fiskebestander enn om den var spesialisert fiskespiser. Kollapser i enkeltbestander antas å ha liten betydning for laksebestandene så lenge andre aktuelle byttedyrbestander er tilgjengelige.
11.6.3 Betydning av fiske på laksens byttedyrbestander
En systematisk gjennomgang av de viktigste byttedyrbestandene til laksen viser at et fåtall av dem er utsatt for fiske. Viktige arter det ikke fiskes på er alle krepsdyrartene, gonatus og mange av fiskeartene, spesielt de mesopelagiske som lysprikkfisk, laksesild og laksetobis. I de fleste studier som finnes på laksediett utgjør disse artene hoveddelen av laksens diett i en eller annen kombinasjon. Arter det fiskes på er f.eks. sild, tobis og kolmule. Sammenbrudd i de aktuelle bestandene pga. fiske vil neppe skje samtidig for flere for bestand, men for et om gangen. Som beskrevet tidligere er det observert totale kollapser i byttedyrbestander (sild) forårsaket i stor grad av fiske med gode lakseår som eneste målbare effekt på laksebestandene.
11.6.4 Kollapser i byttedyrbestandene drevet av klimaforhold
Klimatisk drevne byttedyrbestandssammenbrudd er koblet via hydrografiske/biologiske prosesser til storstilte hydrografiske endringer som påvirker hele næringskjeden. For en topp-predator vil slike sammenbrudd eller reduksjoner i næringskjedens produktivitet kunne få dramatiske effekter fordi mange eller alle de aktuelle næringsdyrenes bestandsnivå kan reduseres parallelt. Samtidig vil den økte konkurransen fra andre arter i nærliggende økologiske nisjer øke relativt. Det forgår for tiden forskning som viser at disse effektene har avgjørende betydning på for eksempel sildebestandens produktivitet i Norskehavet. Kollapser eller kraftige reduksjoner i byttedyrbestander pga. naturgitte forhold, som storstilte endringer i havklima, antas å ha mye større potensiale til å påvirke laksebestandene enn sammenbrudd i byttedyrbestander forårsaket av fiske
11.7 Datagrunnlag og konklusjoner
Datagrunnlaget
Selv om vi har fått atskillig mer kunnskap om laksens liv i havet de senere år, må vi fremdeles konkludere med at kunnskapsnivået er for lavt på mange områder, at våre tidsserier er for korte, og at det vil kreves en betydelig høyning av vår kunnskapsstatus før vi med sikkerhet kan bekrefte (eller avkrefte) gitte vurderinger.
11.7.1 Konklusjoner
11.7.1.1 Havmiljøets betydning
Laksen utgjør en komponent i havets kompliserte økosystem, og er underlagt dynamikken i dette. Vi har påvist at det er en betydelig sammenheng mellom marin dødelighet for laks og havtemperatur, dvs. jo kaldere vann laksen blir eksponert for som postsmolt, jo større dødelighet. Hva som er årsakene til dødeligheten, vet vi ikke, men preliminære analyser av første års vekst av North Esk laks i havet viser en negativ korrelasjon mellom vekst og dødelighet, og en positiv sammenheng mellom vekst og temperatur (Friedland et al. in press). Det er rimelig å anta at lavere vekst vil føre til en høyere predasjonsrate med lavere produktivitet i bestandene som følge.
Forvaltningsmessig er disse resultatene viktige. Fordi det foregår systematiske temperaturmålinger i Atlanterhavet, har man nå fått en mulighet for utvikling av prediktive modeller for overlevelse hos smoltårsklassene. Videre vil en kunne forutsi overlevelse av tosjøvinter laks basert på estimert overlevelse av énsjøvinter laks. Hvor presise slike modeller kan bli, avhenger i stor grad av kvaliteten på de data man legger inn. Kvaliteten kan forbedres ved å få inn bedre informasjon både om viktige miljøparametere og om marin utbredelse, livshistorie, atferd og populasjonsdynamikk hos laks generelt, og for viktige laksepopulasjoner. Etter vår oppfatning synes det som om en utvidelse av temperaturmodellen med bidrag fra forbedret biologisk og økologisk kunnskap om laks, er den mest lovende vei å gå fra dagens ståsted.
Når det marine miljø for laks blir dårligere, blir situasjonen for svake eller sårbare laksepopulasjoner vanskeligere. Modeller for marin overlevelse hos laks kan i fremtiden sette forvaltningen i stand til å sette i verk de nødvendige lokale tiltak på et tidlig tidspunkt.
11.7.1.2 Betydningen av predasjon fra fugl, fisk og sjøpattedyr
I den grad hele eller deler av de senere års negative utvikling i de norske laksebestandene skal kunne forklares ut fra interaksjoner med predatorbestander, må det kunne dokumenteres enten en sterk oppgang i en eller flere av predatorbestandene med derpå følgende øket nedbeiting av laks, eller at annen næringstilgang til en eller flere av predatorbestandene har avtatt kraftig, og at dette næringsbortfallet helt eller delvis er blitt erstattet med laks.
På grunnlag av det tilgjengelige materialet kan det ikke trekkes frem fiskebestander eller grupper av fiskebestander som systematisk kan ha påvirket laksebestandene negativt i den undersøkte perioden og på denne måten ha ført til nedgangen i laksebestandene. Det synes heller som enkelte potensielle predatorbestander er kraftig redusert i denne perioden. Kunnskapene om utviklingen i de aktuelle fiskebestandene er gode, og predasjon fra fisk vurderes ikke som en sannsynlig forklaringsmodell for den negative utviklingen i norske laksebestander etter 1970.
Tilgjengelige data gir ikke mulighet for å bevise at sjøpattedyrene har spilt en rolle i den senere tids negative utvikling i laksebestandene. Det må likevel påpekes at redusert fangst av de fleste arter sjøpattedyr har ført til vekst i bestandene, og at dette kan representere et potensiale for større dødelighet og nedsatt produktivitet i laksebestandene.
Det må her kraftig understrekes at konklusjonene ovenfor ikke bygger på direkte studier av disse forhold, men i stor grad på indirekte vurderinger av potensielle predatorers bestandsnivå og andre forhold. Det ligger her en fare for å undervurdere disse forhold, og da kanskje spesielt den rolle marine sjøpattedyr kan spille for laksebestandenes utvikling. Fordi vi mener at sjøpattedyrene er de eneste predatorene med potensiale til å beite på flersjøvinterlaks i en grad som kan forklare den observerte nedgang i overlevelse hos denne gruppen, anbefaler vi sterkt at sjøpattedyrenes rolle for utviklingen i laksebestandene undersøkes nærmere.
11.7.1.3 Betydningen av konkurranse med andre arter
I havklimatisk kalde og uproduktive perioder observerer vi at store bestander som sild og makrell er magre etter beitesesongen. I disse periodene synes det rimelig at laksebestandene kan oppleve konkurranse fra arter som sild, makrell og kolmule. Disse beiter i de samme havområdene som postsmolten, og det er rimelig å anta at laksebestandene kan oppleve en tilsvarende vekstreduksjon i disse periodene med økt predasjonsdødelighet som følge. Foreløpige analyser foretatt av forfatterne synes å underbygge en slik hypotese, og denne sammenhengen bør utredes nærmere.
11.7.1.4 Betydningen av kollapser i byttedyrbestander
Vi har tidligere konstatert at pga. av laksens opportunistiske ernæringsvaner har fiske på laksens byttedyrbestander liten betydning for laksen så lenge ikke mange bestander reduseres parallelt. Det kan ikke påvises at reduksjoner av byttedyrbestander som følge av fiskerier har påvirket de norske laksebestandene negativt i den aktuelle perioden, tvert imot var det en oppgang i laksebestandene etter kollapsen i bestanden av norsk vårgytende sild. Vi må likevel ta et forbehold om betydningen til loddebestanden i Barentshavet, da det så langt ikke finnes kunnskaper om laksens vandringer i Barentshavet og beitevaner knyttet opp mot lodda i Barentshavet.
Klimatisk drevne endringer i byttedyrbestandene antas å påvirke laksebestandene i større grad enn svingningene som skyldes fiskerier. Det er vist at naturlige svingninger i byttedyrbestandene på grunn av svingninger i havklima fører til store endringer i vekst og kondisjon hos norsk vårgytende sild, som er den største pelagiske bestand i Norskehavet. Slike havklima – svingninger med påfølgende endringer i byttedyrtilgang kan derfor også være en av hovedfaktorene for å forklare svingninger i årsklassestyrke hos norsk laks.
11.7.2 Oppsummering – hovedkonklusjon om havets rolle for de norske laksebestandene.
Storskala havklimatiske svingninger har avspeilet seg og vil fortsatt avspeiles i laksebestandene ved varierende oppgang til elvene. Interaksjoner med predatorbestander, konkurrerende arter eller enkelte byttedyrbestander kan ikke forklare den generelle negative trend en har observert i norske laksebestander fra midten av syttitallet. Vi tar et forbehold om at en mulig vekst i sjøpattedyrbestandene etter sterk tilbakegang eller stopp i fangsten av disse artene, kan ha ført til økt predasjonsdødelighet i laksebestandene. Nye menneskeskapte faktorer er også kommet inn i laksens havfase i den aktuelle perioden og kan ha påvirket laksebestandene negativt både på lokal og global basis.
11.8 Tilrådninger
Forfatterne foreslår følgede tilrådninger i upriorietert rekkefølge:
Man må lære mer om utbredelse og opprinnelse av postsmolt i havet for å identifisere områder hvor postsmolt kan bli tatt som bifangst i pelagiske marine fiskerier og for å undersøke i hvilken grad utbredelsen overlapper med viktige predatorer.
Kontinuitet i havundersøkelsene er viktig da man må etablere tidsserier av fordeling, relativ tetthet, opprinnelse og vekst av postsmolt for å justere/forbedre presisjonsnivået i de prediktive modeller som man er begynt å ta i bruk.
Man bør ta sikte på å videreutvikle metodikken for å estimere mengden postsmolt i Norskehavet.
Man bør ta sikte på å estimere den relative betydning av de forskjellige dødelighets-faktorene for laks i havet.
Man bør prøve å identifisere flere tidsserier av miljørelaterte og biologiske faktorer i laksens utbredelsområde og analysere dem med sikte på å bedre forståelsen av hvordan disse faktorene samvirker i havet.
Det bør merkes mer vill smolt generelt og opprettes flere indeksvassdrag hvor tidsserier av marin overlevelse av smolt kan etableres.
Referanser
Anon. 1983: Report of the working group on north Atlantic salmon. I.C.E.S. C.M. 1983/M.
Anon. 1993. Fagseminar om lakselusproblematikken og tiltaksstrategier. DN-notat (3): 205 pp.
Anon. 1997. Report of the working group on north Atlantic salmon. I.C.E.S. C.M. 1997/Assess:10.
Anon. 1998. Laksen i havet. Resultater fra et forskningsprosjekt ved Færøyene. Tema Nord 1998:520, 63 pp. Nordisk ministerråd, København.
Anon. 1999. Report of the joint ICES/NAFO working group on harp and hooded seals. ICES C.M.1999/ACFM:7.
Crozier, W.W. & Kennedy, G.J.A. 1993. Marine survival of wild and hatchery-reared Atlantic salmon ( Salmo salar L.) from the River Bush, Northern Ireland. Pp. 139–162 in: Mills, D. (ed.). Salmon in the sea and new enhancement strategies. Fishing News Books, Blackwell, Oxford.
Farmer, G.J., Ritter, J.A. & Ashfield, D. 1978. Seawater adaptation and parr-smolt transformation of juvenile Atlantic salmon, Salmo salar . J. Fish. Res. Board Can. 35 : 93–100.
Friedland, K., Reddin, D.G. & Kocik, J.F. 1993. Marine survival of North American and European Atlantic salmon: effects of growth and environment. ICES J. Marine Sci. 50: 481–492.
Friedland, K.D., Hansen, L.P. & Dunkley, D.A. 1998. Marine temperatures experienced by post-smolts and the survival of Atlantic salmon ( Salmo salar L.) in the North Sea area. Fisheries Oceanography, in press.
Friedland, K.D., Hansen, L.P., Dunkley, D.A.& J.C. Maclean. 1998. Linkage between ocean climate, postsmolt growth and survival of Atlantic salmon ( Salmo salar L) in the North Sea area. ICES, J. Marine Sci., in press.
Grimnes, A., and Jakobsen, P.J. 1996. The physiological effects of salmon lice infection on post- smolts of Atlantic salmon. J. Fish Biol. 48: 1179- 1194.
Gunnerød, T.B., Hvidsten, N.A. & Heggberget, T.G. 1988. Open sea releases of Atlantic salmon smolts, Salmo salar L. in Central Norway, 1973–1983. Can. J. Fish. Aquat. Sci., 45, 8: 1340–1345.
Hansen, L.P. 1988. Status of exploitation of Atlantic salmon in Norway. Pp. 143–161 in: Mills, D. & Piggins, D.J. (eds.). Atlantic salmon: Planning for the future. Croom Helm, London.
Hansen, L.P. 1996: The prey of the Atlantic salmon. NASCO Council Meeting, Special session, Gothenburg, Sweden, 13 June 1996. CNL(96)36: 1–17.
Hansen, L.P. & Jonsson, B. 1989. Salmon ranching experiments in the River Imsa: Effect of timing of Atlantic salmon ( Salmo salar ) smolt migration on survival to adults. Aquaculture 82: 367–373.
Hansen, L.P., Jonsson, N. & Jonsson, B.1993. Oceanic migration of homing Atlantic salmon, Salmo salar . Anim. Behav. 45: 927- 941.
Hansen, L.P. & Friedland, K. 1994. Return rates of wild Atlantic salmon tagged as smolts in the River Figgjo, SW Norway 1965–1991 are related to changes in the area of postsmolt habitat. I.C.E.S. North Atlantic salmon Working Group, 3 pp.
Hansen, L.P. & Pethon, P. 1985: The food of Atlantic salmon, Salmo salar L. caught by long–line in Northern Norwegian waters. Journal of Fish Biology 26: 553–562.
Hansen, L.P., Friedland, K.D. & Dunkley, D.A.1995: Examination of survival rates of Atlantic salmon ( Salmo salar L.) from Norway and Scotland and the possible influence of marine habitat area. I.C.E.S. North Atlantic Salmon Working Group, 14 pp.
Hislop, J.R.G. & Shelton, R.G.J. 1993. Marine predators and prey of Atlantic salmon (Salmo salarL.). Pp. 104–118 in: Mills, D. (ed.). Salmon in the sea and new enhancement strategies. Fishing News Books, Blackwell, Oxford.
Holm, M., Holst, J.C.&. Hansen, L.P 1996a: Laks i Norskehavet. Fiskeforsøk og registreringer av laks i Norskehavet og tilgrensende områder juli 1991-august 1995. Fisken og Havet (1): 1–21.
Holm, M., Holst, J.C. and Hansen, L.P. 1996b. Sampling Atlantic salmon in the NE Atlantic during summer: Methods of capture and distribution of catches. ICES C.M. 1996/M12. 7 pp.
Holm, M, Holst, J.C. and Hansen, L.P. 1996c. Distribution of Atlantic salmon smolts in the open sea . Preliminary results of the MARE COGNITUM research programme in the Norwegian Sea and adjacent areas. The Salmon Net, the Magazine of the Salmon Net Fishing Association of Scotland. No XXVII 1996: 27–33.
Holm, M., Huse, I., E. Waatevik, Aure, J.og Døving, K.B. 1984. Følging av utvandrende laksesmolt. I: Atferd hos marine dyr. Foredrag fra symposium, Os. 9.-10. februar 1983: 8–14.
Holm, M., Axelsen, B., Hvidsten, A., Ikonen, E., Johnsen, B.O. and Sturlaugsson, J. 1997. Salmon migration in the Trondheimfjord 1996- Experiences with monitoring acoustically tagged postsmolts with sonar. Second Conference on Fish Telemetry in Europe. La Rochelle, 05- 09. 04.1997. Foredrag, Upubl.
Holst, J. C. & Hvidsten, N. A. 1992. Partrål som prøvetakingsmetode i norsk fiskeriforskning. Fiskets Gang 9/10: 24–26.
Holst, J.C., Hansen, L.P. and. Holm, M. 1996. Observations of abundance, stock composition, body size and food of postsmolts of Atlantic salmon caught with pelagical trawls in the NE Atlantic in summers 1991 and 1995. ICES C.M.1996 M:4. 8 pp.
Holst, J.C., Nilsen, F. Hodneland, K. and Nylund, A. 1993. Observations of the biology and parasites of postsmolt Atlantic salmon, Salmo salar , from the Norwegian sea. Journal of Fish Biology, 42, 962–966.
Hosmer, M.J., Stanley, J.G. & Hatch, R.W. 1979. Effects of hatchery procedures on later return of Atlantic salmon to rivers of Maine. Progr. Fish Cult. 41: 115–119.
Hvidsten, N.A. & Hansen, L.P. 1988. Increased recapture rate of adult Atlantic salmon, Salmo salar L., stocked at high water discharge. J. Fish Biol. 32: 153–154.
Hvidsten, N.A. & Lund, R.A. 1988. Predation on hatchery-reared and wild smolts of Atlantic salmon, Salmo salar L., in the estuary of the river Orkla, Norway. J. Fish Biol. 33: 121–126.
Hvidsten, N.A. & Møkkelgjerd, P.I. 1987. Predation on salmon smolts, Salmo salar , in the estuary of the River Surna, Norway. J. Fish Biol. 30: 273–280.
Hvidsten, N. A., Johnsen, B. O. & Levings, C. D. 1992.Atferd og ernæring hos tvandrende laksesmolt i Trondheimsfjorden. Norsk institutt for naturforskning. Oppdragsmelding 164: 1–14.
Hvidsten, N.A., Johnsen, B.O. & Levings, C.D. 1995. Vandring og ernæring av hos laksesmolt i Trondheimsfjorden og på Frohavet. NINA Oppdragsmelding (332): 1–17.
Håstein, T. & Lindstad, T. 1991. Diseases in wild and cultured salmon: possible interaction. Aquaculture 98: 277–288.
Jacobsen, J.A., Hansen, L.P., Isaksson, A. & Karlsson, L. 1996. Lakseforskning ved Færøyene; et felles nordisk forskningsprosjekt. Fremdriftsrapport til DN og NM 29/5 1996, 8 pp.
Jonsson, N. Hansen, L.P. & Jonsson, B. 1993. Migratory behaviour and growth of hatchery- reared post- smolt Atlantic salmon, Salmo salar . J. Fish Biol.42: 435- 443.
Larsson, P.O. 1977. The importance of time and place of release of salmon and sea trout on the results of stocking. ICES C.M. 1977/M: 42, 4 pp.
Pierce, G.J., Thompson, P.M., Miller, A., Diack, J.S.W., Miller,D. & Boyle, P.R. 1991. Seasonal variation in the diet of the common seals ( Phoca vitulina ) in the Moray Firth area of Scotland. J. Zool. Lond. 223: 641–652.
Poulain, P.M., Warn-Varnas, A. and Niiler, P.P. 1996. Near-surface circulation of the Nordic seas as measured by Lagrangian drifters. Journal of Geophysical Research, 101 (C8)18.237- 18.258.
Reitan, O., Hvidsten, N.A. & Hansen, L.P. 1987. Bird predation on hatchery reared Atlantic salmon smolts, Salmo salar L., released in the River Eira, Norway. Fauna norv. Ser. A . 8: 35–38.
Ritter, J.E. 1989. Marine migration and natural mortality of north American Atlantic salmon ( Salmo salar L.). Can. MS Rep. Fish. Aquat. Sci. 2041, 136 pp.
Rosseland, L. 1971. Fiske av laks i internasjonalt farvann. Jakt- Fiske og Friluftsliv, 100: 190–195; 238–242.
Scarnecchia, D.L. 1984. Climatic and oceanic variations affecting yield of Icelandic stocks of Atlantic salmon ( Salmo salar ). Can J. Fish. Aquat. Sci. 41: 917–935.
Shearer, W.M. 1992. The Atlantic salmon. Natural history, exploitation and future management. Fishing News Books, Blackwell, Oxford, 244 pp.
Shelton, R.G.J., Turrell, W.R., Macdonald, A. 1996. Sampling post-smolt salmon, Salmo salar L., off NW Scotland, June 1996 – Provisional report. The Scottish Office Agriculture and Fisheries Departement, Fisheries Report Services report No 10/96, 17pp.
Shelton, R.G.J., Turrell, W.R., MacDonald, A., McLaren, I.S. and Nicoll, N.T. 1997. Records of post- smoltAtlantic salmon, Salmo salar L., in the Faroe- Shetland Channel in June 1996. Fisheries Research, 31, 159–162.
Skilbrei, O.T., Johnsen, B.O., Heggberget, T.G., Krokan, P.S., Aarset, B. Sagen, T. og Holm, M.. 1998. Havbeite med laks-artsrapport. Norges forskningsråd. 72 pp.
Staurnes, M., Hansen, L.P., Fugelli, K. & Haraldstad, Ø. 1996. Short-term exposure to acid water impairs osmoregulation, seawater tolerance and subsequent marine survival of smolts of Atlantic salmon ( Salmo salar L.). Can. J. Fish. Aquat. Sci. 53: 1695–1704.
Valdemarsen, J. W. and Misund, O. A. 1995. Trawl designs and techniques used by Norwegian research vessels to sample fish in the pelagic zone. In Hylen, A. (ed.). Precision and relevance of pre-recruit studies for fishery management related to fish stocks in the Barents Sea and adjacent waters. Proceeding of the sixth IMR-PINRO Symposium, Bergen, 14–17 June 1994, pp. 129–144.
Wheeler, A. & Gardner, D. 1974. Survey of the literature of marine fish predators on salmon in the Northeast Atlantic. J. Inst. Fish. Mgmt. 5(3): 63–66.
Whelan, K.F. 1993. Decline of sea trout in the west of Ireland: an indication of forthcoming marine problems for salmon. Pp. 171–183 in: Mills, D. (ed.). Salmon in the sea and new enhancement strategies. Fishing News Books, Blackwell, Oxford.