4 Hva er biologisk mangfold?
Knerot
Den sto i skogen
under et tre.
Et furutre.
Jorden var naken
og brun.
Det var en vill orkidé.
Den sto og blomstret
elfenbenshvit.
Ensom og skjønn.
Skogbunnen fattig.
Litt gress.
Barnåler. Mosen var grønn.
Blomsten i skogen
fanget meg inn,
etset seg inn.
Kan ikke glemme
dens lys.
Villblomsten bor i mitt sinn.
Dagny Tande Lid
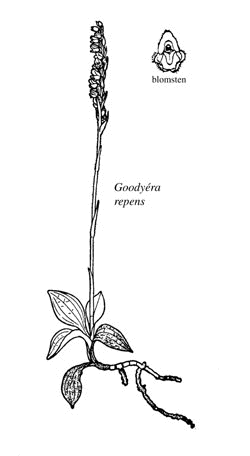
Figur 4.1 Knerot (Goodyéra respen).
Tegning av Dagny Tande Lid
4.1 Begrepsforklaringer
4.1.1 Begrepet biologisk mangfold
Det finnes et særdeles stort antall arter av dyr, planter, sopp og mikroorganismer på jorden. Noe nøyaktig anslag lar seg ikke gi, jf. kap. 4.2.3. I tillegg varierer arveanleggene innenfor den enkelte art mellom de forskjellige populasjonene og individene.
Konvensjonen om biologisk mangfold (heretter kalt biomangfoldkonvensjonen) av 22. mai 1992 definerer i artikkel 2 begrepet biologisk mangfold som ”variasjonen hos levende organismer av alt opphav, herunder bl.a. terrestriske, marine og andre akvatiske økosystemer og de økologiske komplekser som de er en del av; dette omfatter mangfold innenfor artene, på artsnivå og på økosystemnivå”. Definisjonen sikter primært til det som er ”levende”, men fanger også opp samspillet mellom levende organismer og ikke-biologiske faktorer, jf. fremhevelsen av økosystemer. 1
Biologisk mangfold er et komplekst og mangesidig begrep. Det kan inndeles etter ulike dimensjoner som innhold, struktur eller funksjon , se kap. 4.2.2. Mangfoldet kan også betraktes på ulike organisasjonsnivåer, fra gener til populasjoner og arter, og videre til samfunn og økosystemer. I mange praktiske sammenhenger er det imidlertid vanlig å fokusere på artene og deres livsmiljø (habitater). Dels kan det være aktuelt å forvalte artene som bestander eller biosamfunn av mange arter, eller det kan være viktig å sikre artenes livsmiljø. 2 I et helhetsbilde hører også suksesjoner og relasjoner mellom landskap og biologisk mangfold med.
Et viktig aspekt ved bevaring av biologisk mangfold er ivaretakelse av samspill og prosesser , alt fra genetiske prosesser til demografiske prosesser (f.eks. migrasjon, rekruttering, mortalitet, tetthetsregulerende faktorer) og relasjoner mellom arter (f.eks. beiting, predasjon, nedbryting). Et sentralt siktemål er å bevare muligheten for en naturlig dynamikk og evolusjon hos artene.
Mange av de økologiske prosessene er avgjørende for evolusjonære tilpasninger.
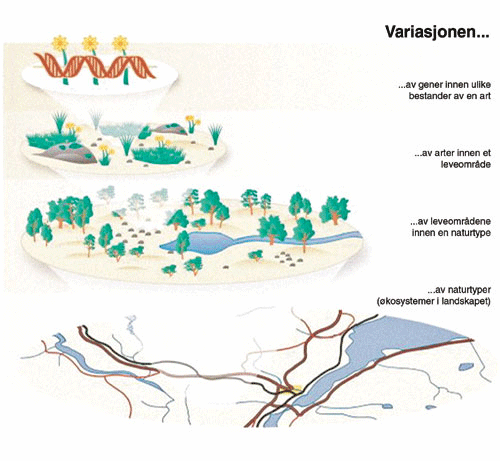
Figur 4.2 Biologisk mangfold (=biomangfold=biodiversitet=livsmangfold): jordens variasjon av livsformer (planter, dyr og mikroorganismer) og deres arvestoff, og samspillet mellom livsformene og det ikke levende miljø. Kan deles inn i mangfold av naturtyper og leveområder, mangfold av arter og genetisk mangfold.
4.1.2 Andre sentrale begreper
Nedenfor gjennomgås en del sentrale begreper som brukes videre i utredningen. Her er det kun de mest grunnleggende begrepene som er omtalt. Betydningen av andre begreper, både av rent naturvitenskapelig og mer forvaltningsmessig karakter, er forklart i ordlisten (vedlegg 3). I tillegg er sentrale begreper i selve lovforslaget definert i utk. § 3.
I dette kapitlet tas det utgangspunkt i biologiske begrepsdefinisjoner. I enkelte tilfeller kan det være et visst avvik mellom biologiske og juridiske definisjoner. For eksempel er definisjonen av ”naturtype” i lovforslaget noe videre enn den biologiske definisjonen av begrepet. De juridiske definisjonene vil bli fortløpende gjennomgått i tilknytning til omtale av utvalgets lovforslag.
4.1.2.1 Det taksonomiske system og begrepet art
Hver art blir betegnet med et toleddet latinsk navn, der første ledd angir slekten og det andre leddet angir arten. Dette er de to siste leddene i det taksonomiske system . Hvis vi bruker vår alminnelige huskatt som eksempel, har vi følgende rekkefølge i det taksonomiske systemet: rike (dyr), rekke ( Chordata – virveldyr), klasse ( Mammalia – pattedyr), orden ( Carnivora – kjøttetere), familie ( Felidae – kattedyr), slekt ( Felis ) og art ( Felissylvestris ).
Arten som enhet er ikke klart definert. Den vanlige inndelingen er morfologisk , det vil si at individer som ser bygningsmessig like ut, antas å tilhøre samme art. Et mer robust kriterium er reproduktiv atskillelse , det vil si at individer av motsatt kjønn som kan få levedyktig og formeringsdyktig avkom, regnes til samme art.
Av åpenbare grunner er kriteriet om reproduktiv adskillelse uegnet for arter med ukjønnet formering. I praksis er det også vanskelig å teste for arter med kjønnet formering, men med forskjellig levested. I stadig større utstrekning benyttes genetiske kriterier (likhet i et utvalg av genetiske markører) for å definere artsgrenser. Sjimpanse og menneske har om lag 95 % felles arvemateriale. De ca. 5 % som er ulikt, er likevel mer enn nok til at sjimpanse og menneske er to selvstendige arter, og vi må sannsynligvis ca. 7 millioner år tilbake for å finne en felles stamform.
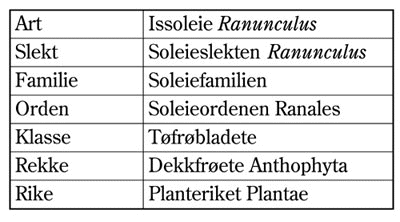
Figur 4.3 Det taksonomiske system og begrepet art, her illustrert ved arten issoleie. Grunnleggeren av dette systemet var den svenske botanikeren Carl von Linné (1707-1778). Han innførte systemet gjennom sitt verk ”Systema naturae” publisert i 1735, mer enn ett århundre før Charles Darwin publiserte sin teori om evolusjonen gjennom boka ”Artenes opprinnelse”.
Enhver art vil over tid gjennomgå evolusjonære endringer. Siden det i de fleste tilfeller er snakk om langsomme endringsprosesser, vil likevel artsbegrepet innen overskuelig fremtid være sentralt i biomangfold-sammenheng. Ettersom enhver art består av populasjoner med større eller mindre grad av genetisk ulikhet, og individer som også er genetisk forskjellige, vil det i mange sammenhenger være mest fruktbart å rette oppmerksomheten mot det totale genetiske mangfoldet innen en art. Dette er ikke minst viktig i bevaringssammenheng, jf. kap. 5.6 og 13.7.4.
4.1.2.2 Planter, dyr, sopp, protister og bakterier
Man regner med fem riker i det taksonomiske systemet: bakterier, protister, sopp, planter og dyr.
Planter kan defineres enten ut fra økologi eller ut fra systematikk. De to måtene å definere planter på dekker nesten det samme, men ikke helt. I økologisk forstand brukes begrepet planter stort sett synonymt med autotrofer eller produsenter, dvs. at de er i stand til selv å bygge opp organisk stoff gjennom fotosyntese eller kjemosyntese. 3 En rekke grupper som regnes til planteriket, viser seg imidlertid å ligge fjernt fra planter flest hvis en ser på faktisk slektskap. Dette gjelder først og fremst de ulike algegruppene utenom grønnalgene, dvs. brunalger (bl.a. tang og tare), rødalger og alle de encellete mer eller mindre gule eller brune algegruppene. 4
I motsetning til planter er dyr heterotrofe, dvs. at de må ha tilførsel av organisk materiale for å skaffe energi, og som stoff til syntese av cellemateriale. Omtrent 75 % av alle beskrevne arter er dyr, og 90–95 % av disse er virvelløse dyr (evertebrater). 5 Virvelløse dyr mangler virvelsøyle eller ryggrad. Hos virveldyr har de fleste artene en virvelsøyle som en del av et kalkskjelett, men hos noen arter består skjelettet av brusk (f.eks. haier og skater). Virveldyrene deles inn i klassene fisk, amfibier, reptiler (krypdyr), fugler og pattedyr.
Begrepene flora og fauna brukes om populasjonen av henholdsvis planter og dyr i et spesielt område. Flora dekker alle planteartene, mens fauna dekker alle dyreartene, både de virvelløse dyrene og virveldyrene. I denne utredningen dekker begrepet flora også sopp, der dette ikke gir grunnlag for misforståelser.
Sopper er heterotrofe organismer som ikke har klorofyll, og de tilhører et eget rike. Soppene er enten saprofytter som ernærer seg av dødt organisk vev, parasitter som ernærer seg av levende organisk vev, mykorrhizapartnere som lever i et gjensidig avhengighetsforhold med høyere planter, 6 eller mellomformer mellom disse. Blant soppene finner vi organismer som har stor betydning i matindustri (gjær, sopp i meieriprodukter) og medisin (penicillin, antibiotika), men også en rekke skadelige arter, primært som parasitter på planter, dyr og mennesker. 7
Protister er en fellesbetegnelse på eukaryote organismer som hovedsakelig opptrer som encellede. Dette er en svært sammensatt gruppe organismer, som omfatter mange uavhengige utviklingslinjer. Blant annet inngår amøber, tøffeldyr, mange frittlevende flagellater og et stort antall forskjellige parasitter.
Bakterier 8 er svært små organismer; bredden varierer fra 0,25 til 2 ìm (1 ìm = 0,001 mm). En bakterie er som oftest én eneste celle som består av plasma uten noen ordentlig kjerne (procaryota). Inne i bakteriene finner vi oftest ett fritt kromosom hvor arveanlegget (DNA) finnes. Bakterier utgjør en svært variabel gruppe som har mange forskjellige funksjoner. Noen er primærprodusenter (f.eks. blågrønnbakterier), noen har kjemosyntese, mange er heterotrofe, og mange er sykdomsorganismer (parasitter). Bakterier er også dominerende aktører i de store biogeokjemiske kretsløpene (f.eks. nitrogen- og karbonsyklusen).
Virus er ikke tatt med i denne oversikten, fordi de definisjonsmessig ikke representerer eget liv. De kan bare formere seg ved hjelp av en vertsorganisme. Virus er partikler med DNA (av og til RNA) i en proteinkappe. De er viktige sykdomsorganismer, og kan ofte regulere forekomst og bestandsstørrelse av andre organismer på alle nivå, inkludert bakterier.
4.1.2.3 Underart, rase, sort, populasjon og bestand
Begrepene underart, rase og sort betegner alle en gruppe individer innenfor en art som har flere felles arvelige egenskaper som gjør at de kan skilles fra andre grupper av individer innenfor arten. Begrepet underart brukes vanligvis når det er tale om ville arter, mens rase og sort stort sett anvendes om grupper innenfor domestiserte arter, henholdsvis om dyr og planter. 9
Begrepet populasjon brukes om en gruppe av individer innen en art som lever innenfor et område til samme tid. Individene som inngår i populasjonen lever mer eller mindre atskilt fra andre individgrupper innen arten. Populasjoner som over tid har levd så vidt atskilt at de skiller seg fra hverandre i frekvensen av visse gener, omtales i noen tilfeller som ulike underarter. Tilsvarende vil populasjoner av arter som på grunn av avl eller foredling har utviklet genetiske særtrekk, bli regnet som egne raser eller sorter. Om ville dyr brukes ofte begrepet bestand i samme betydning som populasjon.
Begrepet bestand brukt om planter betegner en gruppe av planter bestående av et større antall individer, der en enkelt art ofte, men ikke nødvendigvis, er dominerende. Begrepet er mye brukt innen skogbruket, f.eks. ”granbestand”, ”løvtrebestand”, osv.
4.1.2.4 Ville og domestiserte arter
Med en vill art forstås i denne utredningen en art som primært er viltlevende i naturen, herunder i natur som er påvirket av mennesker. Et tilleggskriterium er at arten evolusjonsmessig har fått utvikle seg uten menneskers påvirkning gjennom kultivering eller avl. Dette betyr at arter som er knyttet til f.eks. kulturlandskap eller bymiljø, vil være å regne som ville med mindre de har vært gjenstand for slik direkte menneskelig påvirkning, mens kultiverte arter som har forvillet seg og nå finnes i mer naturpregete omgivelser, ikke regnes som ville arter.
Med domestisering forstår vi den prosessen (kultiveringen eller avlsarbeidet) som fjerner plantene eller dyrene fra det naturlige utvalget over generasjoner. I stedet kultiveres eller avles det målrettet på visse ønskede egenskaper. En domestisert art er en art som har vært gjenstand for seleksjon over så lang tid at genetiske særtrekk er utviklet, og en domestisert populasjon skiller seg genetisk i større eller mindre grad fra viltlevende populasjoner. Det er ikke tilstrekkelig til å anse en art eller populasjon som domestisert at individer holdes i fangenskap eller plantes inn i en hage. Slike individer vil i utgangspunktet fortsatt være å betrakte som ville dyr eller planter.
De artene som først og fremst oppfattes som domestiserte arter, er de som i dagligtale omtales som husdyr og kulturplanter. Samtidig må vi i fremtiden regne med å få en rekke domestiserte bestander av arter som vi i dag regner som ville. Blant annet pågår det arbeid med domestisering av marine arter som f.eks. torsk ( Gadus morhua ), hyse ( Melanogrammus aeglefinus ) og kveite ( Hippoglossus hippoglossus ), og terrestriske arter som f.eks. hjort ( Cervus elaphus ).
I evolusjonsmessig perspektiv kan det være flytende overganger mellom ville og domestiserte populasjoner av arter. For eksempel vil forskjellige former for høsting kunne virke så vidt forstyrrende på den naturlige seleksjonen at artens fremtidige utvikling påvirkes. Denne form for påvirkning endrer likevel ikke på definisjonen av arten som ”vill”. Betydningen av slik menneskelig påvirkning er nærmere utdypet i kap. 4.3.
4.1.2.5 Økosystem, naturtype og vegetasjonstype
Begrepet økosystem henspiller på et sted eller område. Begrepet kan brukes om et større, mer eller mindre ensartet areal (f.eks. et skogområde), en naturtype eller et naturobjekt (f.eks. et tre) som utgjør levested for en rekke organismer. Samtidig omfatter begrepet samspillet mellom organismene på stedet eller i området, og forholdet mellom de biotiske (organismene) og de abiotiske (ikke-biologiske) faktorene som klima, lys, vannhusholdning, næringssalter mv. Dette samspillet omfatter en rekke økologiske prosesser som bidrar til å forme og utvikle det biologiske mangfoldet.
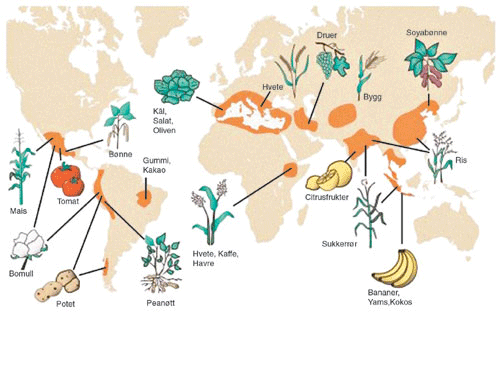
Figur 4.4 Alle kulturplanter stammer fra en art som finnes vill i naturen. Kartet viser de opprinnelige leveområdene for ville planter som i dag utgjør genressursene for våre viktigste matplanter.
Kilde: Primack, R.B. 1993. Essentials of Conservation Biology. Sinauer Associates, Sunderland, Mass.
I biomangfoldkonvensjonen er dette sammenfattet i følgende definisjon av økosystem: et dynamisk kompleks av planter, dyr og mikroorganismer og det ikke-levende miljø rundt dem, som gjennom et samspill utgjør en funksjonell enhet.
En naturtype er en ensartet, avgrenset enhet i naturen som omfatter alt plante- og dyreliv, samt biotiske og abiotiske miljøfaktorer. Variasjonen naturtyper imellom er viktig i seg selv, idet ulike naturtyper gir levesteder for ulike samfunn av dyr, planter og andre organismer.
I likhet med begrepet økosystem, jf. ovenfor, kan også begrepet naturtype anvendes på ulike nivåer. Direktoratet for naturforvaltning (DN) benytter i sin håndbok for kartlegging av biologisk mangfold begrepet ”hovednaturtype” om kategorier som igjen deles inn i naturtyper. 10 Eksempler på slike hovednaturtyper er myr og skog. Eksempler på naturtyper under myr er rikmyr og palsmyr, under skog rik edellauvskog og kystgranskog, se kap. 5.3.
Vegetasjonen er viktig i avgrensningen av naturtyper, og i en del tilfeller beskrives naturtypen bare ut fra vegetasjonen. Begrepet naturtype må likevel ikke forveksles med vegetasjonstype , som kun sier noe om forhold knyttet til plantene. Vegetasjonstype er en klassifiseringsenhet for plantebestand eller plantesamfunn som oppfyller visse fellestrekk. Et plantebestand er som nevnt foran et areal med mer eller mindre jevn fordeling av planter av samme art eller av ulike arter som lever sammen i et fellesskap. Eksempler på vegetasjonstyper er blåbærgranskog, porsfattigmyr og dvergbjørkhei. Vegetasjonstypene karakteriseres av fysisk utforming (vegetasjonssjikt og annen struktur), artssammensetning og mengdefordeling mellom artene. 11 En naturtype vil ofte romme flere vegetasjonstyper. For eksempel kan det innenfor naturtypen deltaområder finnes vegetasjonstyper av både vannvegetasjon, havstrand og skog (f.eks. gråorskog).
Boks 4.1 viser ulike former for avgrensning av naturtyper. Kort sagt bidrar både vegetasjonsutforming, geologiske og kvartærgeologiske forhold, hydromorfologiske forhold, klimatiske forhold og ytre påvirkning (kulturpåvirkning, brann) til å karakterisere de ulike naturtypene. De nevnte forholdene påvirker også dyrelivet, slik at man ved å kjenne naturtypen(e) på et gitt areal, samtidig vil få indikasjoner på hvilket dyreliv som finnes der.
Boks 4.1 Avgrensninger av naturtyper
Eksempler på naturtyper som avgrenses ut fra vegetasjonen, er "bjørkeskog med høgstauder", "gråor-heggeskog" og "stortareskog". "Kystgranskog" avgrenses hovedsakelig ut fra lavsamfunn på trærne. Andre kriterier som kan være sentrale i avgrensningen av naturtyper, er hydromorfologiske forhold og dannelsesmåte - typisk for myrtyper. For palsmyr er i tillegg klimatiske forhold et kriterium, idet denne myrtypen kun opptrer i nordboreal og alpin sone i de mest kontinentale områdene. For naturtyper innen hovednaturtypen kulturlandskap er kulturpåvirkningen et sentralt kriterium i tillegg til vegetasjonen. "Viktige bekkedrag" er eksempel på en naturtype med et landskapsøkologisk utgangspunkt og anvendes som kartleggingsenhet primært i kulturlandskapet. Naturtypen "kroksjøer, flomdammer og meandrerende elveparti" defineres ut fra opphav og flompåvirkning og kan inkludere flere vegetasjonstyper. Avgrensningen av "bekkekløfter" har heller ikke noen direkte kobling til vegetasjonstyper, samtidig som denne naturtypen vil kunne ha et mangfold av vegetasjonstyper på et begrenset areal. For "dammer" og "naturlig fisketomme innsjøer og tjern" er det selve dammen, den fisketomme innsjøen eller tjernet som er utgangspunktet, ikke vegetasjonen. Som navnet "brannfelt" tilsier, er brannpåvirkning et kriterium for denne naturtypen.

Figur 4.5 Leira i Nannestad, Akershus. Typisk meanderende elv. Naturtypen ”kroksjøer, flomdammer og meandrerende elveparti” defineres ut fra opphav og flompåvirkning og kan inkludere flere vegetasjonstyper.
Foto: Øystein Søbye/NN/Samfoto
4.2 Biologisk mangfold på ulike nivåer
4.2.1 Innledning
Det er vanlig å snakke om biologisk mangfold på tre ulike organisasjonsnivåer; økosystemnivå, artsnivå og gennivå, selv om det er mulig å dele inn i flere nivåer.
4.2.2 Økosystemnivå
Det biologiske mangfoldet varierer i innhold (f.eks. hvilke arter som finnes) og struktur (f.eks. hvor store bestandene av artene er) fra ett økosystem til et annet. For eksempel vil forekomsten av treslag som vokser naturlig på et sted, avhenge av klima, topografi, bergart, jordbunnsforhold, vannhusholdning og beiting, samt treslagenes utbredelse og krav til voksested. Det er derfor ikke tilfeldig at treslagssammensetningen er en annen i barskog enn den vi finner på åkerholmer i kulturlandskapet, eller at vegetasjonen er rikere i kalkrike områder enn i andre. Variasjonen kan være stor også innenfor en og samme økosystemkategori, f.eks. fra et våtmarksområde til et annet.
Boks 4.2 Naturlig suksesjon
På dyrket mark som blir liggende brakk, vil det til å begynne med komme opp en mengde ettårige plantearter. Deretter kommer flerårige planter og stauder før lyskrevende småbusker og løvtrær – ofte osp ( Populus tremula ), or ( Alnus spp. ) og bjørk ( Betula spp. ) – gjør sitt inntog. Til slutt kommer skyggetålende treslag, slik som f.eks. gran ( Picea abies ). Innvandringen vil imidlertid avhenge av lokale forhold (klima mv.). Hvilke dyrearter som er i et område vil også avhenge av vegetasjonen og suksesjonsstadiet.
Treslag som først okkuperer et treløst område, kalles pionertreslag, og disse vil over tid avløses av såkalte klimakstreslag. Pionertreslagene er lyskrevende, har rask ungdomsvekst og god formeringsevne. Klimakstreslag tåler skygge og har ofte mer utholdende vekst enn pionertreslagene. Enkelte treslag, som ask ( Fraxinus excelsior ), er skyggetålende i ungdommen, men lyskrevende i moden alder.1
Det er vanlig å dele begrepet suksesjon i to. Primærsuksesjon beskriver den langsiktige kolonisering og utvikling f.eks. etter istider, og dannelse av nytt land ved vulkansk aktivitet. Sekundærsuksesjon skjer på kortere tidsskala etter skogbrann, hogst, opphør av slått osv.
1 Levende Skog 1998a s. 442.
Det forekommer også store variasjoner i økosystemenes funksjoner og produktivitet . Et økosystems funksjoner gjenspeiler de økologiske prosessene på stedet, slik som vannhusholdning, næringsomsetning, kulturpåvirkning og interaksjoner mellom naturlig forekommende arter. Økosystemer kan således fungere som vannreservoar, næringsleverandør, beiteressurs (for så vel ville som tamme dyr) mv., avhengig av karakter og omfang av de økologiske prosessene som finner sted. Produktiviteten gir på sin side uttrykk for produsert biomasse pr. arealenhet, dvs. det kvantitative utbyttet av prosessene. Indikatorer på produktivitet kan f.eks. være mengden av beiteplanter eller næringsdyr.
Økosystemene er dynamiske , det vil si at de hele tiden er gjenstand for naturlige endringsprosesser. Økologiske suksesjoner og lokale ”katastrofer” som brann, insektangrep, stormer mv. er blant de faktorene som gjør at naturen ikke er i en statisk tilstand.
Begrepet suksesjon brukes vanligvis om en langsiktig og retningsbestemt forandring av et økosystem. Gjennom naturlige økologiske suksesjoner gjennomgår økosystemene en serie utviklingsstadier, der nye plante- og dyresamfunn avløser hverandre etter hvert som organismene skaper nye miljøbetingelser. Naturlig suksesjon kan illustreres med det som skjer når dyrket mark blir liggende brakk, se boks 4.2.
Generelt er diversiteten høyest et stykke ut i et suksesjonsforløp, men avtar som regel mot klimaks som ofte preges av noen få dominerende arter. Økosystemene påvirkes også av menneskelig aktivitet og av globale prosesser som f.eks. klimaendringer. Disse påvirkningsfaktorene kan gripe inn i økosystemenes naturlige dynamikk og endre det biologiske mangfoldet på økosystemnivå så vel som på artsnivå og gennivå. Menneskeskapte påvirkningsfaktorer er nærmere omtalt i kap. 7.2.
4.2.3 Artsnivå
Mangfoldet på artsnivå kommer til uttrykk gjennom antall arter og variasjonen i individtall mellom arter. Så langt er færre enn to millioner arter vitenskapelig beskrevet og gitt navn på verdensbasis, mens det årlig klassifiseres i størrelsesorden 15 000 nye arter. 12 Anslagene på uoppdagete arter – hovedsakelig fisk, sopp, insekter og mikroorganismer – varierer fra fem millioner til mer enn hundre millioner. Fortsatt oppdages og beskrives også ”nye” pattedyr- og plantearter hvert år.
Trolig utgjør encellede organismer det største mangfoldet både artsmessig og genetisk. Dette er imidlertid en gruppe som er lite kjent, og hvor vanlige systematiske kriterier for artsbestemmelse er problematiske. Samtidig er dette en gruppe med stor betydning både for økosystemfunksjon og stoffsyklus, og som potensielt utnyttbare organismer i mange sammenhenger. På den annen side rommer denne gruppen også en rekke sykdomsfremkallende organismer.
Mangfoldet av arter er ikke konstant, idet noen arter dør ut og nye arter dannes. Artsdannelse er først og fremst et resultat av geografisk isolasjon. Når isolasjonen opphører, vil individer fra den tidligere isolerte bestanden krysse seg med individer fra de opprinnelige bestandene og få fruktbart avkom. Ved lang tids isolasjon kan den isolerte bestanden bli så ulik den opprinnelige arten at de ikke lenger kan gi fruktbare hybrider, og artsdannelsen er da et faktum. Den gradvise utviklingen fra en til flere atskilte arter er belyst bl.a. i studier av fluesnappere, se boks 4.3.
Menneskers aktivitet kan også påvirke artsdannelsen. Dette kan f.eks. skje ved flytting av arter til nye geografiske områder eller ved produksjon av nye, ofte mellomliggende, habitater. To eksempler som kan illustrere dette er nydannelse av margeritter på Kanariøyene og nydannelse av arter innen kurvplanteslekten geitskjegg, se boks 4.3.
Boks 4.3 Eksempler på artsdannelse
Fluesnappere
DNA-analyser av fluesnappere tyder på at det opprinnelig fantes én fluesnapperart. Under istiden ble den opprinnelige bestanden antakelig isolert i tre forskjellige refugier – det vil si midlertidige tilfluktssteder; et på Balkan-halvøya, et i Italia og et på Iberiahalvøya. På disse store halvøyene holdt skogen stand under istiden. Bestandene var isolert over så lang tid at de utviklet seg forskjellig, og resultatet ble tre arter: Balkan-fluesnapper (Ficedula semitorquata ), svarthvit fluesnapper ( Ficedula hypoleuca ) og halsbåndfluesnapper ( Ficedula albicollis ). Etter istiden har to av bestandene ekspandert nordover, og i Mellom-Europa kom halsbåndfluesnapper og svarthvit fluesnapper sammen i en hybridsone. Hvis de parrer seg med hverandre, får de en hybrid – altså en blanding av to arter – med nedsatt fruktbarhet. Siden hybridene er delvis fruktbare, var ikke artsdannelsen hos fluesnapperne helt fullendt. Likevel kan artsdannelsen fortsette når de er ført sammen igjen. Det er påvist at omtrent to prosent av de aktuelle hunnene velger feil partner. Naturen ”straffer” disse gjennom hybridavkom med nedsatt fruktbarhet, mens de hunnene som velger partner av samme art premieres. Det antas at dette på sikt vil føre til en fullstendig artsdannelse.1
1 Hammersmark 1998.
Margeritter
På Kanariøyene finnes det en rekke margerittarter som er tilpasset ulike voksesteder og som fra naturens side er godt atskilt. I åpne, tørre, lavereliggende områder vokser bl.a. en smalbladet margeritt med middels store blomster ( Argyrantheum frutescens ). Høyere opp, i den fuktige laurbærskogen nordvest på Tenerife vokser en bredbladet storblomstret art ( A. broussonetii ). På noen lokaliteter i mellomliggende høydenivå er det bygget vei og delvis avskoget. Her har de to artene møttes og gitt opphav til to nye arter, ( A. lemsii ) og ( A. sundingii ).
Geitskjegg
Kurvplanteslekten geitskjegg ( Tragopogon ) har tre arter som opprinnelig var hjemmehørende i Europa. Hybrider mellom de ulike artene er kjent herfra, men de er ikke fertile. På 1900-tallet ble artene brakt med mennesker til Nord-Amerika, der de i visse områder ble ganske vanlige ugress. Med utgangspunkt i disse artene ble det dannet to fertile hybrider som er genetisk konstante og anerkjent som egne arter, T. mirus (T. dubius x porrifolius) og T. miscellus (T. dubius x pratensis) . Innen andre slekter er lignende nydannelse av arter kjent, f.eks. mellom europeiske og nordamerikanske arter. Menneskets flytting av arter har i disse tilfellene ført til ny artsdannelse og økt biologisk mangfold.
4.2.4 Gennivå
Selv om mangfoldet er stort på artsnivå, gir ikke artsantallet som sådant noe fullstendig bilde av det biologiske mangfoldet. Innen en og samme art er det genetisk variasjon både mellom individene og mellom forskjellige populasjoner. Denne variasjonen må også tas i betraktning for at bildet av mangfoldet skal bli komplett. Et mye omtalt eksempel på genetisk variasjon innen en art er det store mangfoldet av laksestammer i Norge. Fordi villaksen finner tilbake til sin egen fødeelv for å gyte, har det utviklet seg en rekke forskjellige stammer av laks ( Salmo salar ), hver genetisk og lokalt tilpasset livet i sin elv.
Det er miljøfaktorene – både de abiotiske og de biotiske – som bestemmer hvilke egenskaper hos organismene som gir best mulighet til å overleve i et bestemt miljø. Seleksjon av visse genutgaver innen en art kan etter hvert skape organismer som er godt tilpasset de lokale miljøforhold. Isolasjonsmekanismer er således en nødvendig forutsetning for lokal genetisk tilpasning, og det finnes en rekke slike mekanismer som hindrer eller reduserer flyten av genmateriale mellom arter og mellom de enkelte bestandene. Fysiske barrierer som medfører hel eller delvis geografisk isolasjon mellom bestander, er en effektiv isolasjonsmekanisme. Laksens vandring tilbake til elven hvor den vokste opp, er en annen isolasjonsmekanisme, der atferden bremser flyten av genmateriale mellom bestander. 13 Nettopp laksens hjemvandring er et godt eksempel på at en lokal tilpasning til de lokale miljøforhold er viktig, og ”genetisk forurensning” fra rømt oppdrettslaks anses for et betydelig problem, se kap. 7.2.4.6.
Dersom det er snakk om svært små bestander, kan isolasjon være negativt. Små og isolerte bestander har ofte liten genetisk variasjon, noe som gir så vel økt risiko for innavl som redusert mulighet for kombinasjon av nye genvarianter – og med det evolusjon. Det er med andre ord den genetiske variasjonen som danner grunnlag for videre tilpasning og utvikling. I tråd med dette er genetisk variasjon et element som tillegges større vekt i tenkningen omkring bevaring av biologisk mangfold nå enn tidligere, og det er også av samme grunn viktig å bevare bestander av en viss størrelse.
Det er grunn til å tro at biologisk mangfold i økende omfang vil bli beskrevet med utgangspunkt i genetisk variasjon, i takt med at gener og basesekvenser 14 hos artene i norsk flora og fauna kartlegges i stadig større omfang. Ny kunnskap har påvist at artsgrensene i mange tilfeller er uklare. De er oftest basert på enkle morfologiske egenskaper, f.eks. farger, som kan forklares med et par mutasjoner. Spesielt gjelder dette for lavere organismer. For mange organismer, spesielt de encellede, vil genetiske analyser være den eneste muligheten for å fastslå diversitet og taksonomisk tilhørighet.
Også for høyerestående organismer er den genetiske variasjonen en viktig indikator for å fastslå opprinnelse, spredningsmønstre mv. Blant annet har svenske undersøkelser vist at den sør-skandinaviske ulven ( Canis lupus ) skiller seg genetisk fra dyrepark-ulv. 15
I 1960-årene regnet forskerne med at den sørskandinaviske ulvestammen var utryddet, men tidlig i 1980-årene ble det igjen observert ulveyngling i grenseområdene mellom det sørøstlige Norge og Sverige. Deretter holdt bestanden seg på rundt ti individer, alle etterkommere av det ene paret som etablerte seg her. Det vil si at det var en betydelig innavl. Alle var nære slektninger, og den genetiske variasjonen var svært lav. Studier viste at innavl resulterte i medfødte misdannelser, bl.a. arvelig blindhet. Fra 1991 fikk plutselig flokken ny livskraft og begynte å øke raskt, og det ble etablert 10–11 mindre flokker som spredte seg til nye områder. Genetiske studier av denne ulvebestanden har vist at denne økningen skjedde etter at en enslig ulv vandret inn fra Nord-Finland eller Russland. Analysene viser at denne enslige innvandreren brakte et nytt genetisk materiale inn i den lille, innavlete bestanden. 16 Dette viser hvordan genflyt kan være av svært stor betydning for små, isolerte bestander.
4.3 Naturgitt, kulturpåvirket og kulturbetinget mangfold
I omtale av biologisk mangfold skilles det ofte mellom det naturgitte mangfoldet og det kulturpåvirkete eller kulturbetingete mangfoldet. Til naturgitt biologisk mangfold regnes mangfold som ikke er utsatt for menneskers påvirkning i merkbar grad. Mangfoldet kan likevel ha endret seg over tid, som følge av økosystemenes naturlige dynamikk, jf. kap. 4.2.2. At det biologiske mangfoldet i et område anses som naturgitt, behøver ikke å bety at området er urørt i den forstand at det ikke finnes spor etter menneskelig aktivitet. Det kan finnes spor i terrenget i form av fornminner el.l. uten at det biologiske mangfoldet er endret, i alle fall ikke i et omfang som er sporbart i dag.
Kulturpåvirket biologisk mangfold er påvirket av mennesker gjennom en eller flere typer handlinger. Påvirkningen kan ha funnet sted ved f.eks. oppdyrking, hogst eller introduksjon av fremmede arter. Gjennom slike inngrep endres økosystemenes karakter. Graden av endringer varierer, og dersom påvirkningen er midlertidig, vil tidsfaktoren kunne bidra til at endringene ”viskes ut” igjen gjennom suksesjon. Ofte er det vanskelig å trekke noe klart skille mellom naturgitt og kulturpåvirket biologisk mangfold. Det kan derfor være mer hensiktsmessig å betrakte det biologiske mangfoldet på en skala fra naturgitt – via ulike grader av påvirket – til kraftig manipulert. Et eksempel som illustrerer denne glidende overgangen mellom det naturgitte og det kulturpåvirkede, er den påvirkningen som forårsakes av ulike former for høsting, se boks 4.4.
Mens de menneskeskapte endringene i noen områder kan ha ført til en utarming av det biologiske mangfoldet, kan de i andre områder ha virket berikende ved å legge til rette for et økt biologisk mangfold. Ikke minst førte tidligere tiders oppdyrking til at en rekke nye arter fikk levelige vilkår også i Norge. Slike arter ble dels innført aktivt av mennesker og dels spredt som følge av forbedrete livsvilkår. Mange av de gamle innførte plantene må i dag kunne sies å inngå i den stedegne flora. Slike arter er gjerne konkurransesvake og representerer ikke generelt noen trussel mot det naturgitte biologiske mangfoldet. 17
Generelt har tidsfaktoren mye å si for hvordan kulturpåvirket natur og biologisk mangfold betraktes. Jo lengre tid som er gått siden en introduksjon av en art eller omforming av et område har funnet sted, jo større er sannsynligheten for at arten vil bli betraktet som hjemlig eller de nye landskapselementene som integrert og hjemmehørende i deler av området. For eksempel ble knoppsvane ( Cygnus olor ) introdusert til norsk natur på 1800-tallet, og arten oppfattes nok – på lik linje med de gamle innførte plantene i jordbrukslandskapet – av mange som stedegen, selv om den strengt vitenskapelig ikke er del av Norges naturgitte biologiske mangfold.
Boks 4.4 Eksempler på påvirkning av biologisk mangfold gjennom jakt og fangst
En rekke arter som vi er vant til å tenke på som del av det naturgitte biologiske mangfoldet, er mer eller mindre påvirket av menneskers seleksjon gjennom jakt og fangst. I enkelte elgbestander skjer opptil 85 til 90 % av uttaket gjennom jakt, og da sier det seg selv at dette er en faktor av vesentlig betydning for den evolusjonsmessige utviklingen hos elg ( Alces alces ). Rettet avskyting, der man styrer jakten mot bestemte kjønns- og alderskategorier av dyr, kan føre til store endringer i bestandens sammensetning og vekstrate, sett i forhold til hva som ville ha vært situasjonen uten slik jakt.
Resultatene fra en undersøkelse på Vega i Nordland viser at en reduksjon i oksealder og -andel i en elgbestand som følge av rettet avskyting vil kunne ha demografiske konsekvenser for bestanden. Okseandelen på Vega er blitt for lav til at alle elgkyr blir bedekt til riktig tid, noe som har ført til forsinket kalving med påfølgende lave kalvevekter, åringsvekter og mulig redusert fruktbarhet. Videre synes kalvekjønnsraten å dreie mot færre oksekalver i en bestand med hovedsakelig unge okser. Dette vil i sin tur påvirke bestandens vekstrate1.
Hos grågåsa ( Anser anser ) er det påvist en utvikling med stadig tidligere hekking. Til en viss grad kan dette skyldes klimaendringer, men en like sannsynlig forklaring er at seleksjon gjennom jakt påvirker gjessenes hekking. De gjessene som først flyr sørover når jakten starter, er de ikke-hekkende individene, samt kull som er blitt klekket ekstra tidlig. Disse unngår dermed å bli skutt under jakten, og jakten blir en seleksjonsmekanisme som favoriserer de individene som hekker tidlig og trekker tidlig. Denne mekanismen blir sterkere jo tidligere jakten starter.2
Det er fastsatt minstemål for stort sett alle arter fisk i norsk fiske, jf. forskrift 10. oktober 1989 nr. 1095 om maskevidde m.v. ved fangst av fisk. Slike minstemål er satt for å forbedre rekrutteringen til de eldre årsklassene og dermed sikre en bærekraftig ressursutnyttelse. Samtidig vil en overrepresentasjon av store, livskraftige individer i uttaket kunne ha betydning for bestandens videre utvikling. Tiltak som anses effektive for å hindre overbeskatning, kan dermed vise seg å ha andre, evolusjonsmessige virkninger på lang sikt. For eksempel er minstemålet for hummer ( Homarus gammarus ) satt til 24 cm, noe som beskytter førstegangsgytende hunner, mens de eldre hunnene blir gjenstand for fangst. De førstegangsgytende hunnene har imidlertid gjennomgående færre egg med dårligere kvalitet enn større hunner, med de konsekvenser dette kan ha for bestandsutviklingen.
1 Sæther et al. 2001.
2 http://www.jaktogfiske.info
I Sverige defineres en innført art som naturlig forekommende dersom den enten er innført passivt før år 1900 og deretter har reprodusert spontant, eller er innført aktivt før år 1800 og deretter har utviklet lokale tilpasninger (uten foredling), se boks 13.9.
Mens det er glidende overganger mellom naturgitt og kultur påvirket biologisk mangfold, er det et klarere skille mellom naturgitt og kultur betinget biologisk mangfold. Kulturbetinget biologisk mangfold er ikke bare påvirket av mennesker, det er i tillegg avhengig av slik påvirkning over tid for å kunne opprettholdes. Et typisk eksempel er kulturlandskapstyper som har oppstått som følge av driftsformer i landbruket. Når ekstensive driftsformer erstattes med nye, mer intensive driftsformer, vil disse kulturlandskapstypene lett kunne bli skadelidende.
Menneskers påvirkning på naturen kan også endre forholdet mellom arter som forekommer naturlig. Dette skjer bl.a. ved at menneskelig aktivitet endrer abiotiske faktorer slik som lysforhold, vannhusholdning etc, slik at man – bevisst eller ubevisst – favoriserer enkelte arter på bekostning av andre. Utbredelsen av en art kan således være kulturbetinget, selv om artens tilstedeværelse i Norge ikke er det.
4.4 Forholdet til genmodifiserte organismer
Mennesker har i tusener av år foredlet planter og dyr for å få frem best mulig egenskaper. Ved hjelp av genteknologi kan egenskaper endres uten å avle over mange generasjoner. Genteknologi gjør det mulig å sette arvestoff (DNA) sammen på nye måter og overføre DNA mellom organismer (planter, dyr, mikroorganismer). Genmodifiseringen kan bestå i at organismen får ekstra gener, at gener blir forandret eller at deler av eller hele gener fjernes. Genmodifiserte organismer lages nå innenfor mange felter: mat, fôr, tekstiler, legemidler osv. 18
Hvis genmodifiserte organismer blir en del av miljøet, er det en viss fare for at de kan overføre de nye genene som de har fått, til liknende organismer. For planter kan spredning skje via pollen, og for dyr kan krysning med ikke-modifiserte dyr føre til at de nye genene spres. Innsatte gener eller DNA-fragmenter kan være ustabile, og det er en risiko for at de fremmede genene overføres til andre arter ved at f.eks. bakterier tar opp i seg DNA fra de modifiserte organismene og fører dette videre til helt andre arter ved en mekanisme som kalles horisontal genoverføring. Når gener kombineres på nye måter, kan det være vanskelig å forutsi hvordan de vil fungere. Det er også en mulighet for at modifiserte arter i visse tilfeller kan fortrenge naturlige arter eller at de har andre utilsiktede virkninger på miljøet.
Fotnoter
Om begrepet økosystem, se kap. 4.1.2.5.
Blom et al. 2004 med henvisning til Noss 1990.
Det finnes planter som ikke er autotrofe (f.eks. enkelte orkideer). Soldogg er autotrof, men tar opp nitrogen via insekter.
http://www.toyen.uio.no
http://www.miljolare.no
Mykorrhizasoppene har stor positiv innvirkning på plantenes vekst og vitalitet. Mykorrhiza er en planterot infisert med slike sopper. Soppene bedrer næringsopptaket hos plantene og beskytter røttene mot sykdom og miljøstress. Soppene på sin side nyter godt av næringsforbindelser som trærne produserer gjennom fotosyntesen.
http://www.nisk.no
Bakteriene deles inn i to grupper: eubakterier og archaebakterier.
Om ville og domestiserte arter, se kap. 4.1.2.4.
Direktoratet for naturforvaltning 1999a.
Fremstad & Moen (red.) 2001 s. 229 til 231.
http://news.nationalgeographic.com
NOU 1999:9 vedlegg 4.
DNA fra alle levende organismer er satt sammen av de fire basene adenin (A), cytosin (C), guanin (G) og thymin (T). Basene danner par A-T og G-C som danner en dobbelttrådet DNA-helix. Sekvensen av baser i DNA inneholder instruksjonen om hvordan en organisme blir laget. Antall basepar varierer fra noen få tusen for enkle virus til milliarder basepar for komplekse organismer.
Ellegren, Savolainen & Rosén 1996
Vila et al. 2003
Direktoratet for naturforvaltning 1999b s. 1 til 3.
http://www.bion.no